Nấm Pucciniomycetes gây bệnh cây trồng
Teleomorph, anamorph, pycniospores, aeciospores, urediniospores, teliospores, basidiospores, Puccinia, Uromyces, Hemileia, Gymnosporangium, Phakopsora, Coleosporium

Thuộc ngành Nấm Đảm (Basidiomycota), Pucciniomycetes đại diện cho một trong những nhóm sinh vật gây bệnh thực vật phức tạp, tinh vi và quan trọng nhất về mặt kinh tế trên quy mô toàn cầu, thường được biết đến với tên gọi phổ biến là nấm rỉ sắt (rust fungi).
Ước tính lớp này chứa khoảng 8.400 loài đã được mô tả, chiếm hơn 8% tổng số loài nấm đã biết và khoảng một phần ba số lượng nấm đảm. Sự đa dạng này không chỉ thể hiện ở số lượng loài mà còn ở phạm vi ký chủ rộng lớn, từ các loài dương xỉ cổ đại, cây lá kim (gymnosperms) đến các loài thực vật có hoa (angiosperms) hiện đại.
Đặc điểm cấu tạo
Thể hữu tính (teleomorph)
Teleomorph ở Pucciniomycetes bắt đầu khi hình thành telium và teliospore, sau đó phát triển thành basidium, nơi xảy ra meiosis để tạo basidiospore.
Telium là cơ quan sinh bào tử của nấm, nơi tạo ra teliospore, loại bào tử dày vỏ, đóng vai trò trung tâm của giai đoạn hữu tính (teleomorph) trong vòng đời của Pucciniomycetes. Telium thường xuất hiện dưới dạng đốm hoặc ổ màu nâu đen trên lá, thân hoặc bẹ lá của cây, bề mặt có thể phẳng, nổi u, hoặc vỡ bung ra để lộ bào tử. Telium là giai đoạn cực kỳ quan trọng giúp nấm sống qua mùa đông hoặc điều kiện bất lợi và là nơi bắt đầu giai đoạn hữu tính.
Teliospore (bào tử hậu, bào tử đông) là bào tử dày vỏ của nấm, được sinh ra trong telium và đóng vai trò then chốt trong giai đoạn hữu tính (teleomorph) của vòng đời. Teliospore có thành rất dày, màu nâu hoặc đen giúp nấm sống sót qua mùa đông, hạn, lạnh. Teliospore thường gồm 2 tế bào (bicellular), nhưng tùy loài có thể 1 tế bào, mỗi tế bào chứa 2 nhân (dikaryotic) trước khi nảy mầm.
Teliospore không gây bệnh cho cây mà chỉ dùng để chuyển sang giai đoạn hữu tính.
Basidium là cơ quan sinh sản hữu tính đặc trưng của Nấm Đảm (Basidiomycota), nơi diễn ra giảm phân (meiosis) để tạo ra basidiospore, tức bào tử hữu tính của Nấm Đảm. Basidium ở Pucciniomycetes có dạng ống mảnh, mọc ra từ teliospore, chia thành 3-4 tế bào ngang gọi là phragmobasidium. Mỗi tế bào phragmobasidium sinh 1 basidiospore.
Basidiospore là bào tử hữu tính của Nấm Đảm (Basidiomycota), là sản phẩm cuối cùng của quá trình giảm phân trong basidium. Basidiospore có kích thước nhỏ, thường 10-20 µm, hình cầu, trứng, giọt nước hoặc elip (tùy loài). Thành bào tử mỏng, màu vàng nhạt hoặc trong suốt, không dày và sậm như teliospore. Basidiospore mang 1 nhân haploid (n) hình thành bên ngoài basidium (exogenous spore).
Chu trình phát triển của thể hữu tính (teleomorph) ở nấm Pucciniomycetes là một quá trình sinh học phức tạp, đánh dấu sự chuyển đổi từ giai đoạn sợi nấm song nhân (n+n) sang giai đoạn lưỡng bội (2n), sau đó giảm phân để trở về đơn bội (n). Khi cây chủ già đi hoặc thời tiết chuyển lạnh, nấm ngừng sản sinh bào tử vô tính (urediniospore) và chuyển sang tạo bào tử đông (teliospore). Từ hệ sợi nấm song nhân (n+n) trên cây chủ, các teliospore vách dày được hình thành. Hợp nhân (karyogamy) là bước quan trọng nhất. Hai nhân đơn bội (n+n) trong mỗi tế bào của teliospore sẽ hợp nhất lại thành một nhân lưỡng bội (2n). Sau khi hợp nhân, teliospore bước vào trạng thái ngủ nghỉ, vách tế bào trở nên rất dày và sẫm màu (hóa melanin) để bảo vệ nhân lưỡng bội bên trong khỏi lạnh giá và tia cực tím. Giai đoạn này không có hoạt động phân chia tế bào, chỉ duy trì sự sống tiềm sinh. Khi thời tiết ấm lên và độ ẩm tăng cao, teliospore kết thúc trạng thái ngủ và bắt đầu nảy mầm. Đây là lúc thể hữu tính phát triển mạnh mẽ nhất. Mỗi tế bào của teliospore nảy mầm, mọc ra một ống dài gọi là basidium. Nhân lưỡng bội (2n) từ trong teliospore di chuyển vào basidium, thực hiện giảm phân để tạo ra 4 nhân đơn bội (n). Các vách ngăn ngang xuất hiện, tạo thành 4 tế bào con, mỗi tế bào chứa 1 nhân đơn bội (phragmobasidium). Từ mỗi tế bào phragmobasidium mọc ra một cuống nhỏ, nhọn, đầu mỗi cuống phình ra tạo thành bào tử đảm (basidiospore). Nhân đơn bội từ phragmobasidium di chuyển qua cuống để chui vào trong đảm bào tử. Khi chín, đảm bào tử được phóng thích cưỡng bức (bắn ra) vào không khí nhờ cơ chế giọt nước để bay đi tìm vật chủ mới.
Một điểm đặc biệt trong chu trình này là bào tử đảm (Basidiospore) không thể lây nhiễm lại vật chủ cũ (nơi sinh ra teliospore).
Thể vô tính (Anamorph)
Giai đoạn vô tính của Pucciniomycetes được định danh là giai đoạn Uredinial. Đây là giai đoạn duy nhất trong vòng đời có khả năng tự lặp lại liên tục, tạo nên các đợt dịch rỉ sắt tàn phá mùa màng.
Uredinium là cơ quan/bộ phận sinh bào tử vô tính của nấm gỉ, nơi tạo ra urediniospore. Uredinium hình thành dưới lớp biểu bì của lá, thân hoặc bẹ lá. Ban đầu xuất hiện như khối mô nhỏ, màu vàng nhạt, nằm ẩn dưới biểu bì. Khi trưởng thành đẩy phồng và làm nứt biểu bì để lộ ra bột bào tử màu vàng cam, vàng nâu hoặc nâu đỏ.
Urediniospore (bào tử hạ) là bào tử vô tính quan trọng nhất của Pucciniomycetes.
Đây là tác nhân gây nhiễm chính trong suốt mùa sinh trưởng của cây, làm bệnh lan nhanh theo chu kỳ ngắn. Tùy loài, nhưng hình dạng thường là hình cầu, hình elip, hoặc hơi tròn, đường kính khoảng 18-35 µm, bề mặt có gai nhỏ (echinulate) đặc trưng. Màu vàng cam, vàng nâu hoặc nâu đỏ đặc trưng của bệnh rỉ sắt. Thành dày vừa, không dày như teliospore, có khoảng 2-8 lỗ nảy mầm (germ pores). Nhân dikaryotic (n+n), hai nhân luôn tồn tại song song trong cùng một tế bào bào tử.
Khi sợi nấm song nhân (n+n) đã khai thác đủ dinh dưỡng từ các tế bào nhu mô lá xung quanh, nấm nhận tín hiệu (thường do mật độ sợi nấm cao hoặc thay đổi sinh lý cây chủ) để ngừng mọc lan và bắt đầu tập trung lại. Các sợi nấm di chuyển và tập trung dày đặc ngay bên dưới lớp biểu bì (epidermis) của cây. Chúng đan xen chặt chẽ tạo thành một lớp đệm mô nấm gọi là primordium (khởi nguyên thể). Tại lớp đệm này, các tế bào nấm bắt đầu phân chia theo hướng thẳng đứng (vuông góc với bề mặt lá) và biệt hóa thành hai phần rõ rệt:
- Tế bào chân (basal cells) nằm dưới cùng, làm nhiệm vụ neo giữ và cung cấp dinh dưỡng.
- Tế bào sinh bào tử (sporogenous cells) nằm phía trên, các tế bào này bắt đầu vươn dài ra để chuẩn bị tạo bào tử.
Tế bào sinh bào tử phân chia, tạo ra một tế bào dài, mảnh ở phía dưới gọi là cuống (pedicel), ngắn và trong suốt. Phần đầu của tế bào phình to ra, vách tế bào dày lên nhanh chóng. Các hạt sắc tố (carotenoid - tạo màu rỉ sắt) và chất dinh dưỡng được bơm vào phần phình to này. Cặp nhân song nhân ($n+n$) phân chia đồng bộ. Một cặp nhân nằm lại ở cuống (hoặc tiêu biến), cặp nhân còn lại di chuyển vào phần đầu phình to để tạo thành bào tử hạ (Urediniospore) hoàn chỉnh.
Khi hàng ngàn bào tử hạ được hình thành đồng thời và lớn lên, chúng tạo ra một áp lực trương (turgor pressure) khổng lồ đẩy lên lớp biểu bì của lá cây. Lớp biểu bì cây chủ không chịu nổi áp lực sẽ bị xé toạc. Các mảnh biểu bì bị rách thường dựng đứng lên bao quanh ổ nấm, tạo thành một cổ áo (epidermal collarette) đặc trưng mà ta có thể quan sát. Khối bào tử dạng bột màu vàng/nâu lộ ra ngoài không khí, sẵn sàng bị gió cuốn đi.
Bào tử hạ (n+n) từ cây bệnh bay theo gió và rơi xuống lá cây khỏe. Khi có sương hoặc giọt nước, bào tử nảy mầm mọc ra ống mầm (germ tube). Ống mầm bò trên bề mặt lá cho đến khi gặp khí khổng (stomata). Tại đây, đầu ống mầm phình to tạo thành đĩa bám (appressorium) để neo chặt. Từ đĩa bám, một kim xâm nhập (infection peg) chọc thủng khe khí khổng để chui vào bên trong mô lá. Sợi nấm phát triển len lỏi giữa các tế bào thực vật (gian bào). Chúng đâm các vòi hút (haustoria) vào bên trong tế bào cây chủ để hút đường và chất dinh dưỡng nuôi nấm. Sau khi tích lũy đủ dinh dưỡng (khoảng 1-2 tuần), sợi nấm tập trung lại dưới biểu bì và hình thành Uredinium mới. Hàng ngàn Urediniospore mới được sinh ra, phá vỡ biểu bì lá và bay đi để bắt đầu một vòng lặp lây nhiễm mới.
Khi nguồn dinh dưỡng từ cây chủ cạn kiệt hoặc thời tiết chuyển lạnh, chu trình vô tính sẽ dừng lại. Thay vì tạo ra bào tử hạ (màu vàng/nâu), tơ nấm sẽ chuyển sang sản xuất bào tử đông (Teliospore) màu đen. Đây chính là lúc nấm chuyển từ giai đoạn vô tính (Anamorph) sang giai đoạn chuẩn bị cho hữu tính (Teleomorph).
Tính đa hình thái bào tử
Tính đa hình thái bào tử (Pleomorphism) là đặc điểm nổi bật nhất và phức tạp nhất của lớp nấm Pucciniomycetes. Một loài nấm rỉ sắt điển hình (nấm có chu trình sống đầy đủ - Macrocyclic) có thể sản sinh tới 5 loại bào tử khác nhau trong vòng đời của nó, được đánh số từ 0 đến IV theo quy ước quốc tế.
Giai đoạn 0: Bào tử phấn (Spermatia/Pycniospores) là giai đoạn khởi đầu cho quá trình sinh sản hữu tính. Chúng có hình dạng giống chiếc bình (flask-shaped), thường nằm ở mặt trên của lá cây vật chủ trung gian với trạng thái nhân đơn bội (n). Bào tử rất nhỏ, không thể tự nảy mầm gây bệnh. Túi tinh tiết ra mật ngọt để dụ côn trùng mang bào tử phấn (n) sang các sợi nấm tiếp nhận (receptive hyphae) của dòng khác giới tính để thực hiện quá trình phối chất (plasmogamy), tạo ra hệ sợi nấm song nhân (n+n), tạo ra sự đa dạng di truyền và tiền đề cho Giai đoạn I.
Giai đoạn I: Bào tử xuân (Aeciospores) là kết quả trực tiếp của quá trình thụ tinh ở giai đoạn 0. Chúng thường mọc ở mặt dưới của lá (đối diện với vị trí túi tinh), có hình dạng giống chiếc cốc (cup-shaped) hoặc chuông với trạng thái song nhân (n+n). Bào tử hình chuỗi, thường có màu vàng cam rực rỡ. Bào tử xuân là bào tử chuyển chủ, không thể nhiễm lại cây chủ nơi chúng sinh ra mà bay theo gió để lây nhiễm sang cây chủ chính.
Giai đoạn II: Bào tử hạ (Urediniospores) là giai đoạn sinh sản vô tính bùng nổ như đã đề cập ở phần trước.
Giai đoạn III: Bào tử đông (Teliospores) là giai đoạn nấm khi kết thúc mùa vụ hoặc gặp điều kiện bất lợi. Chúng có vách rất dày, sẫm màu (đen/nâu đen), thường có 2 tế bào. Trạng thái nhân ban đầu là (n+n), sau đó hợp nhân thành lưỡng bội (2n) nhờ quá trình hợp nhân (karyogamy).
Giai đoạn IV: Bào tử đảm (Basidiospores) là giai đoạn nảy mầm và tái tổ hợp di truyền như đã đề cập ở phần trước. Bào tử đảm bay đi tìm vật chủ trung gian để bắt đầu lại Giai đoạn 0.
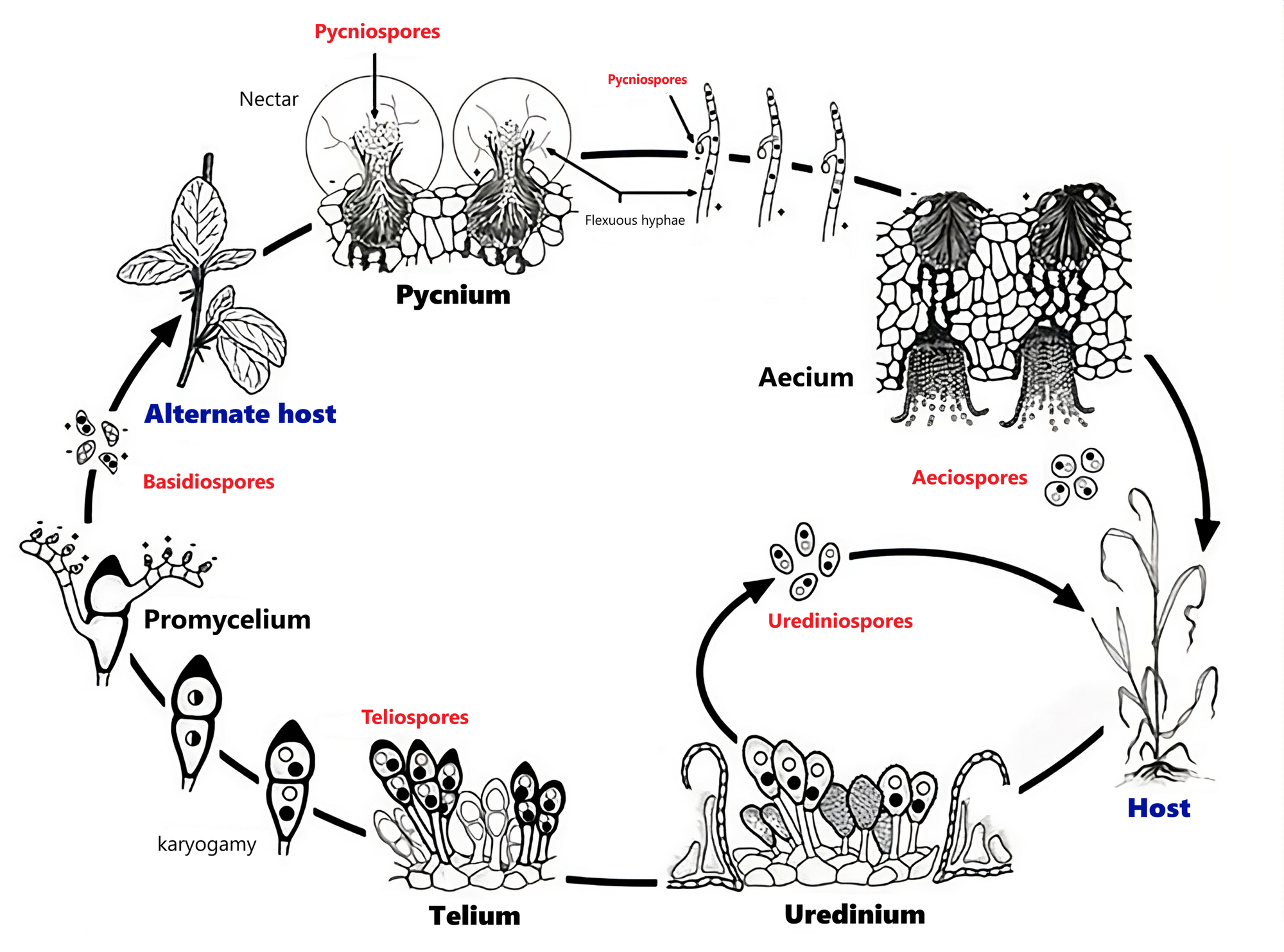
Vòng đời sinh trưởng, phát triển
Dựa trên số lượng các giai đoạn bào tử xuất hiện trong vòng đời, các loài nấm rỉ sắt được phân loại thành:
- Vòng đời lớn (Macrocyclic): Sở hữu đầy đủ cả 5 giai đoạn bào tử (0, I, II, III, IV). Đây là dạng tiến hóa hoàn chỉnh nhất.
- Vòng đời bán khuyết (Demicyclic): Thiếu giai đoạn bào tử hạ (II). Nấm chuyển trực tiếp từ giai đoạn bào tử xuân (I) sang bào tử đông (III).
- Vòng đời nhỏ (Microcyclic): Vòng đời rút ngắn tối đa, chỉ còn bào tử đông (III) và bào tử đảm (IV). Bào tử đông có thể nảy mầm ngay sau khi hình thành mà không cần giai đoạn nghỉ.
Sự sinh trưởng và phát triển của Pucciniomycetes phụ thuộc chặt chẽ vào nhiệt độ và độ ẩm, những yếu tố quyết định khả năng nảy mầm và xâm nhiễm. Mỗi loài có ngưỡng nhiệt độ tối ưu riêng. Hầu hết bào tử nấm rỉ sắt (đặc biệt là bào tử hạ) cần một màng nước tự do trên bề mặt lá (sương, mưa) trong khoảng 6-12 giờ liên tục để nảy mầm.
Các chi nấm bệnh trọng yếu
Chi Puccinia
Đây là Chi lớn và quan trọng nhất, gây nhiều bệnh rỉ sắt nghiêm trọng trên cây trồng. Đặc điểm hình thái quan trọng để phân biệt Puccinia là bào tử đông (teliospores) có vách ngăn ngang chia làm 2 tế bào, có cuống rõ ràng, không rụng, thường có màu nâu đậm hoặc đen. Mỗi tế bào có một lỗ nảy mầm.
Chi Uromyces
Về mặt hình thái và di truyền, Uromyces rất gần gũi với Puccinia. Điểm khác biệt duy nhất là bào từ đông chỉ có 1 tế bào (unicellular), có cuống rõ ràng, thường có núm dày ở đỉnh bào tử.
Chi Hemileia
Chi này đặc biệt quan trọng đối với cây cà phê. Bào tử hạ có hình dạng quả thận (kidney-shaped), mặt lưng có gai sần sùi, mặt bụng trơn nhẵn. Bào tử đông (Teliospore) có hình cầu hoặc góc cạnh, trơn, nảy mầm ngay lập tức (không qua giai đoạn ngủ nghỉ), mọc ra từ khí khổng (suprastomatal sori). Các cụm bào tử nhô ra qua lỗ khí khổng ở mặt dưới lá, tạo thành lớp bột phấn màu vàng cam. Mặt trên lá tương ứng xuất hiện các đốm vàng dầu loang lổ.
Chi Gymnosporangium
Chi này gây bệnh rỉ sắt trên cây họ Táo và họ Hoa hồng. Bào tử đông có cấu trúc 2 tế bào (tương tự Puccinia). Khi gặp ẩm, cuống bào tử (pedicel) ngậm nước và trương phồng lên tạo thành các khối nhầy giống thạch (gelatinous), màu cam hoặc vàng cam rất nổi bật trên thân cây.
Chi Phakopsora
Chi này nổi tiếng với các bệnh rỉ sắt gây hại nghiêm trọng trên cây họ Đậu, đặc biệt là đậu tương. Bào tử đông có cấu trúc đa bào không theo quy luật cố định, sắp xếp thành nhiều lớp chồng lên nhau, không có cuống (sessile). Tạo thành các lớp vỏ (crusts) màu nâu đen dính chặt dưới biểu bì lá, rất khó tách rời để quan sát từng bào tử riêng lẻ. Bào tử hạ thường có gai (echinulate) và lỗ nảy mầm phân bố rải rác. Đây là giai đoạn thường gặp nhất trên đồng ruộng.
Chi Coleosporium
Chi Coleosporium là một chi nấm rỉ sắt rất đặc biệt và dễ nhận biết, thường gây ra các bệnh rỉ sắt có màu cam tươi sáng (khác với màu nâu rỉ sắt sẫm của Puccinia). Bào tử đông ban đầu là đơn bào nhưng khi chín bào tử không nảy mầm tạo ra ống mầm bên ngoài, mà vách ngăn sẽ hình thành ngay bên trong bào tử, chia bào tử thành 4 ngăn, không có cuống riêng lẻ. Các bào tử xếp xít nhau tạo thành một lớp vỏ (crust) hoặc lớp sáp (waxy layer) phẳng, dính chặt dưới biểu bì lá. Bào tử hạ mọc thành chuỗi (catenate). Giữa các bào tử trong chuỗi thường có các tế bào đệm nhỏ, dễ gãy để giúp phát tán bào tử.
Các Chi nấm rỉ sắt khác như Chrysomyxa, Coleosporium, Cronartium thường gây bệnh cho cây hạt trần hoặc cây trồng cảnh.
Quản lý hóa học và chiến lược kiểm soát bền vững
Biện pháp hóa học thường là tuyến phòng thủ cần thiết khi áp lực bệnh cao hoặc khi điều kiện thời tiết thuận lợi cho nấm phát triển (ẩm độ cao, nhiệt độ ấm). Việc quản lý các bệnh do Pucciniomycetes gây ra phụ thuộc rất nhiều vào các biện pháp kiểm soát hóa học. Việc hiểu rõ các nhóm thuốc và cơ chế tác động (MoA) của chúng là điều kiện tiên quyết để quản lý hiệu quả và bền vững. Các nhóm chính được sử dụng để kiểm soát Pucciniomycetes bao gồm: Triazoles (FRAC 3), Strobilurins (FRAC 11), Carboxamides (FRAC 7) và các hoạt chất chuyên biệt...
Xu hướng quản lý hóa học hiện đại đang dịch chuyển khỏi việc sử dụng các hoạt chất đơn lẻ và tiến tới việc sử dụng các hỗn hợp được nghiên cứu kỹ lưỡng. Các sản phẩm thuốc trừ nấm hiện đại thường được bào chế sẵn dưới dạng hỗn hợp của hai hoặc nhiều hoạt chất có cơ chế tác động khác nhau, thể hiện tác dụng hiệp đồng (synergistic effects).
Tính kháng thuốc trừ nấm là mối đe dọa lớn nhất đối với tính bền vững của nền nông nghiệp hiện đại. Các mầm bệnh Pucciniomycetes, với khả năng tái tổ hợp di truyền nhanh và biến đổi cao, đặc biệt dễ phát triển tính kháng. Để ngăn chặn quá trình này, các nguyên tắc quản lý tính kháng bắt buộc phải được áp dụng:
- Không bao giờ sử dụng cùng một nhóm thuốc (cùng nhóm FRAC/MoA) lặp lại trong nhiều lần phun. Phải luân phiên với các nhóm thuốc có MoA hoàn toàn khác.
- Ưu tiên sử dụng các sản phẩm đã được trộn sẵn 2 hoặc nhiều MoA. Lý do là xác suất để một bào tử nấm có đột biến kháng cả hai MoA cùng một lúc là cực kỳ thấp.
- Cần giới hạn số lần phun tối đa mỗi mùa vụ.
- Tích hợp các loại thuốc tiếp xúc, đa điểm (Nhóm M) vào chương trình phun. Các thuốc này tác động lên nhiều điểm trong tế bào nấm, khiến khả năng phát triển tính kháng gần như bằng không.
Sử dụng các giống kháng bệnh (nếu có), quản lý nước tưới và bón phân cân đối, thực hành vệ sinh đồng ruộng nghiêm ngặt, thu gom và tiêu hủy tàn dư cây bệnh để giảm thiểu nguồn bệnh sơ cấp.