Oomycetes gây bệnh cây trồng
Water molds, zoospores, oospores, Phytophthora, Pythium, Plasmopara, Peronospora, Pseudoperonospora, Bremia, Peronosclerospora, Albugo, Aphanomyces

Oomycetes thường được gọi là nấm mốc nước (water molds) hay nấm trứng, đại diện cho một nhóm vi sinh vật nhân thực (eukaryotic) dị dưỡng có tầm quan trọng đặc biệt trong bệnh học thực vật và nông nghiệp toàn cầu. Trong lịch sử nghiên cứu nấm học, Oomycetes đã từng được phân loại vào giới Nấm (Fungi) trong một thời gian dài. Sự phân loại này dựa trên các đặc điểm hình thái tương đồng mang tính hội tụ (convergent evolution), bao gồm kiểu sinh trưởng dạng sợi (filamentous growth), cấu trúc hệ sợi nấm (mycelium), và phương thức dinh dưỡng hấp thu (absorptive nutrition) thông qua việc tiết enzyme ngoại bào để phân giải cơ chất.
Tuy nhiên, cuộc cách mạng trong sinh học phân tử và di truyền học vào cuối thế kỷ 20 đã lật ngược quan điểm này. Các phân tích phát sinh chủng loại (phylogenetic analysis) dựa trên trình tự gene RNA ribosome (rRNA) và các protein bảo thủ đã xác nhận rằng Oomycetes không có quan hệ họ hàng gần gũi với nấm thật (Eumycota). Thay vào đó, chúng thuộc về giới Stramenopila, đặt Oomycetes vào cùng nhóm tiến hóa với tảo nâu (brown algae). Phát hiện này không chỉ mang tính hàn lâm mà còn giải thích tại sao các chiến lược kiểm soát nấm bệnh truyền thống thường thất bại khi áp dụng cho Oomycetes, do sự khác biệt căn bản về sinh hóa và cấu trúc tế bào.
Đặc điểm cấu tạo
Sự phân biệt giữa Oomycetes và Nấm thật là nền tảng cốt lõi để hiểu về cơ chế gây bệnh và cơ chế tác động của thuốc bảo vệ thực vật. Sự khác biệt này thể hiện ở mọi cấp độ từ cấu trúc siêu vi đến sinh hóa.
| Đặc điểm | Oomycetes (Nấm trứng) | Nấm thật (True Fungi) | Ý nghĩa Bệnh học & Quản lý |
|---|---|---|---|
| Vị trí phân loại | Giới Stramenopiles (Chromista) | Giới Fungi | Oomycetes có quan hệ gần gũi với Tảo nâu và Tảo silic hơn là Nấm. |
| Thành phần vách tế bào | Cellulose (β-1,4-glucan) và β-1,3/β-1,6 glucans. Rất ít hoặc không có chitin (< 1%) |
Chitin là thành phần cấu trúc chính | Thuốc ức chế tổng hợp chitin vô hiệu với Oomycetes. Cần dùng thuốc nhóm CAA hoặc Phenylamides. |
| Bộ nhiễm sắc thể | Lưỡng bội (2n) ở giai đoạn sinh dưỡng; giảm phân xảy ra ở cơ quan sinh dục | Đơn bội (n) hoặc song nhân (n+n) trong phần lớn vòng đời | Trạng thái 2n giúp Oomycetes có khả năng thích nghi nhanh và biến dị di truyền cao hơn. |
| Cấu trúc ty thể | Mào ty thể dạng ống (tubular cristae) | Mào ty thể dạng phiến (flattened cristae) | Đặc điểm giải phẫu vi thể phản ánh nguồn gốc tiến hóa khác biệt. |
| Sinh tổng hợp lysine | Con đường diaminopimelic acid (DAP) | Con đường alpha-aminoadipic acid (AAA) | Khác biệt về enzyme chuyển hóa cơ bản. |
| Sterol màng tế bào | Không tự tổng hợp sterol hoặc tổng hợp fucosterol | Ergosterol là sterol chủ đạo | Các thuốc nhóm Triazole (ức chế ergosterol) thường kém hiệu quả với Oomycetes. |
| Bào tử động (zoospores) | Có 2 roi khác nhau (heterokont): 1 roi lông tơ (trước) + 1 roi trơn (sau) |
Thường không có (trừ nấm Chytridiomycota có 1 roi trơn phía sau) |
Zoospores bơi trong nước tự do là đơn vị lây nhiễm chủ yếu trong mùa mưa. |
| Vách ngăn sợi nấm | Sợi nấm cộng bào (coenocytic), hiếm khi có vách ngăn (aseptate) | Sợi nấm có vách ngăn rõ ràng (septate) | Dòng chất nguyên sinh di chuyển tự do giúp vận chuyển dinh dưỡng cực nhanh, gây bùng phát bệnh cấp tính. |
Thành tế bào
Thành tế bào của Oomycetes là một cấu trúc động, đóng vai trò quan trọng trong việc duy trì hình thái tế bào, bảo vệ chống lại áp suất thẩm thấu và là rào cản đầu tiên tương tác với hệ thống miễn dịch của thực vật. Không giống như nấm thật sử dụng mạng lưới chitin cứng chắc, Oomycetes đã tiến hóa để sử dụng cellulose và các β-glucan khác làm khung xương chịu lực. Các nghiên cứu hóa sinh đã chỉ ra rằng cellulose chiếm khoảng 4-20% thành phần vách tế bào, trong khi β-1,3-glucan và β-1,6-glucan chiếm tỷ lệ lớn. Một phát hiện quan trọng trong sinh học phân tử Oomycetes là việc xác định họ gen CesA (Cellulose Synthase) chịu trách nhiệm tổng hợp cellulose.
Hệ sợi nấm cộng bào
Hệ sợi của Oomycetes phát triển dưới dạng các ống hình trụ phân nhánh, và đặc điểm nổi bật nhất là tính chất cộng bào (coenocytic) hay không vách ngăn (aseptate). Trong cấu trúc này, không có các vách ngăn ngang chia sợi nấm thành các tế bào riêng biệt. Thay vào đó, tế bào chất là một khối liên tục chứa hàng ngàn nhân tế bào trôi nổi tự do. Các vách ngăn (septa) chỉ xuất hiện trong hai trường hợp: (1) để cô lập các bộ phận sinh sản (túi bào tử hoặc cơ quan sinh dục) khỏi phần còn lại của sợi nấm, hoặc (2) để ngăn cách các phần sợi nấm già cỗi, bị tổn thương.
Cấu trúc cộng bào mang lại lợi thế sinh học to lớn: dòng tế bào chất (cytoplasmic streaming) có thể vận chuyển năng lượng, enzyme và vật liệu xây dựng thành tế bào từ các phần già của khuẩn lạc đến các đỉnh sinh trưởng (hyphal tips) với tốc độ rất cao. Điều này giải thích tại sao các bệnh do Oomycetes như bệnh mốc sương khoai tây hay bệnh sương mai dưa chuột có thể bùng phát và tiêu hủy toàn bộ cánh đồng chỉ trong vài ngày.
Chuyển hóa
Về mặt sinh lý, Oomycetes thể hiện nhiều đặc điểm của giới Stramenopiles. Chúng tổng hợp amino acid lysine thông qua con đường diaminopimelic acid (DAP), giống như thực vật và vi khuẩn, thay vì con đường alpha-aminoadipic acid (AAA) như nấm thật.
Một đặc điểm sinh lý quan trọng khác liên quan đến quản lý bệnh là sự thiếu hụt sterol. Các chi như Phytophthora và Pythium được gọi là các loài dị dưỡng sterol, nghĩa là chúng không thể tự tổng hợp sterol từ đầu mà phải lấy từ môi trường hoặc vật chủ để sinh sản hữu tính và sinh trưởng tối ưu. Ngược lại, nấm thật phụ thuộc vào ergosterol để duy trì tính toàn vẹn và linh động của màng tế bào. Chính vì vậy, các nhóm thuốc ức chế sinh tổng hợp sterol (như nhóm triazoles, imidazoles ức chế C14-demethylase) thường có hiệu lực rất thấp hoặc không có hiệu lực đối với Oomycetes, buộc ngành bảo vệ thực vật phải tìm kiếm các đích tác động khác như hô hấp ty thể hay tổng hợp lipid màng.
Sinh sản và vòng đời
Khả năng sinh sản của Oomycetes là một kiệt tác của sự thích nghi tiến hóa, cho phép chúng tồn tại qua các điều kiện khắc nghiệt và bùng phát nhanh chóng khi điều kiện thuận lợi.
Sinh sản vô tính là giai đoạn chịu trách nhiệm cho các đợt dịch bệnh bùng phát (epidemics) trong mùa vụ. Cấu trúc trung tâm của quá trình này là Túi bào tử (Sporangium). Túi bào tử được hình thành trên các cành mang túi bào tử (sporangiophores) chuyên biệt. Điểm độc đáo của Oomycetes là khả năng lựa chọn phương thức nảy mầm dựa trên nhiệt độ môi trường - một cơ chế cảm ứng nhiệt tinh vi:
- Nảy mầm trực tiếp ở nhiệt độ cao (thường >15-20°C tùy loài), túi bào tử hoạt động giống như một bào tử nấm thông thường (conidium). Nó mọc ra một ống mầm (germ tube) và xâm nhập trực tiếp vào mô cây.
- Nảy mầm gián tiếp (Indirect germination - Tạo động bào tử): Ở nhiệt độ thấp (mát mẻ, thường <15°C) và có sự hiện diện của nước tự do, tế bào chất bên trong túi bào tử phân chia thành nhiều đơn vị nhỏ (thường từ 8-30), mỗi đơn vị phát triển thành một động bào tử (Zoospore).
Động bào tử là cấu trúc lây nhiễm hiệu quả nhất của Oomycetes trong môi trường ẩm ướt. Chúng là các tế bào trần (không vách), hình quả lê hoặc hình thận, có hai roi. Roi trước dạng lông (tinsel) tạo lực kéo, roi sau dạng trơn (whiplash) đóng vai trò bánh lái. Sự phối hợp nhịp nhàng của hai roi cho phép chúng bơi trong màng nước mỏng trên bề mặt lá hoặc trong nước khe đất. Chúng sở hữu các thụ thể cảm nhận tín hiệu hóa học từ dịch tiết của rễ cây (như đường, amino acid, ethanol) và tín hiệu điện trường từ bề mặt rễ non. Điều này giúp chúng tìm đến đúng vị trí dễ xâm nhập nhất (như vùng chóp rễ hoặc khí khổng). Khi tiếp xúc với bề mặt vật chủ, zoospore rụng roi, tiết ra glycoprotein để bám dính chặt vào bề mặt, và nhanh chóng tổng hợp vách tế bào mới để trở thành bào nang (cyst). Bào nang này sau đó nảy mầm để xâm nhập.
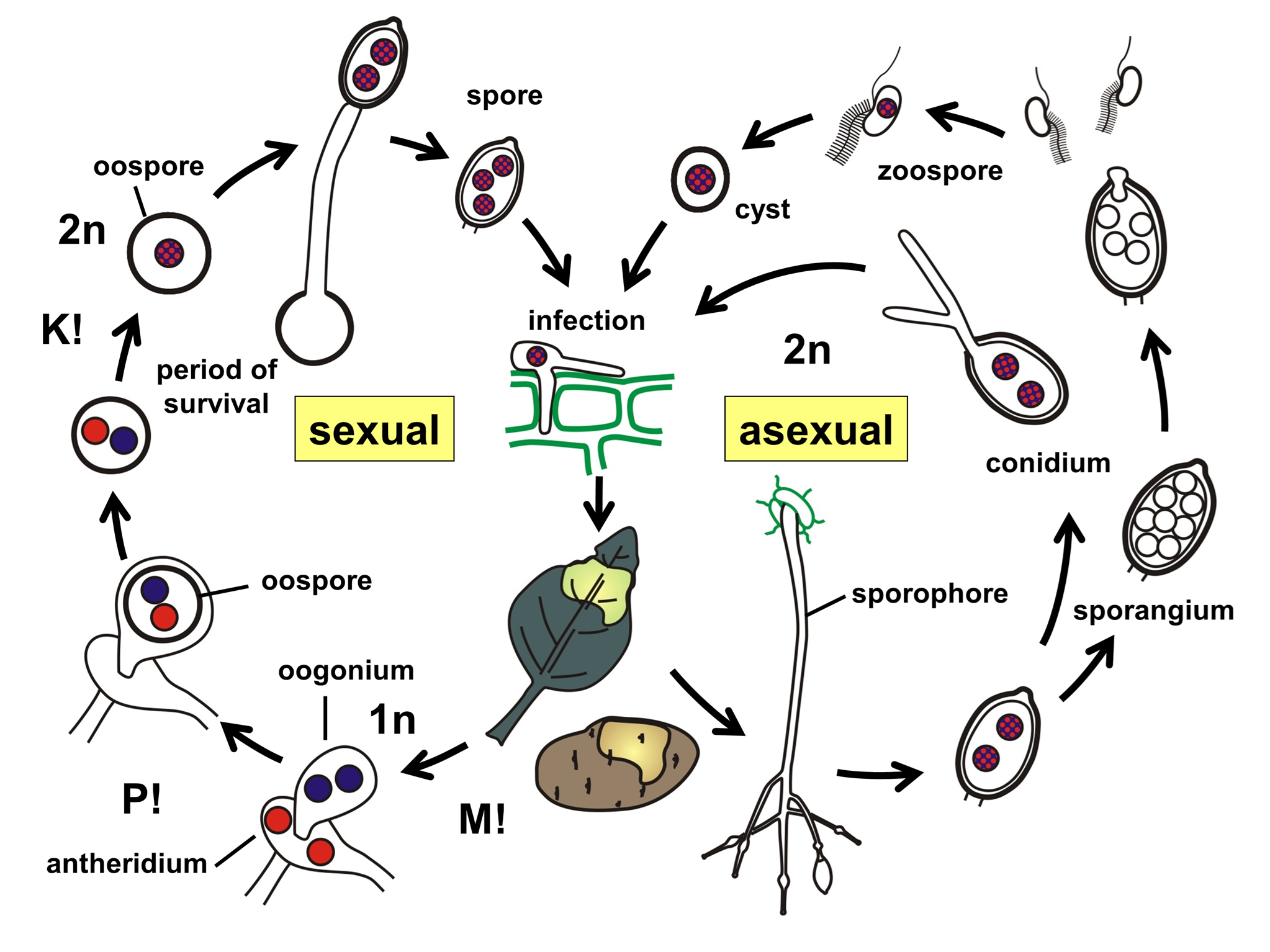
Sinh sản hữu tính thường diễn ra vào cuối mùa vụ hoặc khi điều kiện môi trường trở nên bất lợi (khô hạn, thiếu dinh dưỡng), tạo ra bào tử trứng (oospores). Quá trình thụ tinh diễn ra giữa hai cơ quan giao phối khác biệt về hình thái:
- Túi noãn (oogonium): Cơ quan cái, hình cầu lớn, chứa một hoặc nhiều noãn cầu (oospheres) đơn bội.
- Túi hùng (antheridium): Cơ quan đực, nhỏ hơn, bám vào túi noãn. Nhân từ túi hùng di chuyển qua ống thụ tinh (fertilization tube) vào túi noãn để thụ tinh cho noãn cầu, tạo thành hợp tử lưỡng bội, sau đó phát triển thành oospore.
Oospore có vách tế bào dày, nhiều lớp, chịu được khô hạn, lạnh giá và sự phân hủy của vi sinh vật đất. Chúng có thể tồn tại trong đất từ vài tháng đến nhiều năm (thậm chí 10 năm với Pythium). Khi điều kiện thuận lợi quay trở lại, oospore nảy mầm tạo ra túi bào tử hoặc ống mầm để khởi đầu một chu kỳ bệnh mới (primary inoculum).
Cơ chế xâm nhiễm
Sau khi bám dính lên bề mặt cây trồng, Oomycetes sử dụng các chiến lược xâm nhập khác nhau tùy thuộc vào loài và vị trí tấn công.
Xâm nhập trực tiếp (direct penetration) phổ biến ở các loài gây bệnh rễ và một số bệnh lá. Sau khi đóng kén, bào tử nảy mầm tạo ống mầm. Tại đầu ống mầm hình thành một cấu trúc phình to gọi là giác bám (appressorium). Giác bám tạo ra áp lực cơ học cực lớn kết hợp với việc tiết ra một hỗn hợp các enzyme phân giải (Cell Wall Degrading Enzymes - CWDEs) như cutinase, cellulase, pectinase để khoan thủng lớp cutin và vách tế bào biểu bì. Một kim xâm nhập (penetration peg) mọc ra từ giác bám đâm xuyên qua vách tế bào để đưa sợi nấm vào bên trong.
Xâm nhập qua khí khổng (stomatal penetration) đặc trưng của nhóm nấm sương mai và rỉ trắng. Ống mầm của các loài này định hướng phát triển về phía khí khổng. Khi đến nơi, chúng phình to tạo thành cấu trúc xâm nhập (substomatal vesicle) trong khoang dưới khí khổng, từ đó phát triển các sợi nấm lan vào mô thịt lá (mesophyll). Chiến lược này giúp chúng tránh phải phá vỡ lớp cutin dày, nhưng đòi hỏi cơ chế nhận biết vị trí khí khổng cực kỳ chính xác.
Dựa vào cách thức lấy dinh dưỡng từ vật chủ, Oomycetes được chia thành ba nhóm chính:
- Ký sinh chuyên bắt buộc (obligate biotrophs) với đại diện: Plasmopara, Bremia, Peronospora, Albugo. Chúng chỉ có thể sống và sinh sản trên mô cây còn sống. Chúng phát triển sợi nấm len lỏi giữa các khoảng gian bào (intercellular) và đưa các cấu trúc chuyên biệt gọi là vòi hút (haustoria) đâm xuyên qua vách tế bào vào bên trong (nhưng không phá vỡ màng sinh chất). Haustoria đóng vai trò như một cổng giao tiếp, hút dinh dưỡng từ tế bào chất của cây đồng thời tiết ra các protein effector để ức chế phản ứng tự vệ của tế bào.
- Ký sinh bán chuyên (hemibiotrophs) với đại diện là Phytophthora. Giai đoạn đầu, chúng hành xử như ký sinh chuyên bắt buộc (biotrophic phase), sống âm thầm và lấy dinh dưỡng mà không giết chết tế bào. Sau đó, chúng chuyển sang giai đoạn hoại sinh (necrotrophic phase), tiết ra độc tố và enzyme phá hủy tế bào hàng loạt để tiêu thụ chất dinh dưỡng từ mô chết, gây ra các vết thối rữa đặc trưng.
- Hoại sinh (necrotrophs) với đại diện là Pythium.Chúng tấn công thô bạo ngay từ đầu, tiết ra một lượng lớn enzyme phân giải pectin (pectinolytic enzymes) và độc tố để giết chết tế bào nhanh chóng. Mục tiêu là biến mô cây thành một khối dịch lỏng dinh dưỡng để hấp thu. Đây là nguyên nhân gây ra triệu chứng thối nhũn (damping-off) ở cây con.
Ở cấp độ phân tử, sự tương tác giữa Oomycetes và thực vật tuân theo mô hình zig-zag của hệ thống miễn dịch.
- PTI (PAMP-Triggered Immunity): Khi Oomycetes tấn công, vách tế bào của chúng giải phóng các phân tử đặc trưng gọi là PAMPs (Pathogen-Associated Molecular Patterns), ví dụ như β-glucans, Pep-13, hoặc các enzyme transglutaminase. Các thụ thể trên màng tế bào thực vật nhận diện PAMPs và kích hoạt phản ứng miễn dịch cơ bản (PTI), bao gồm việc củng cố vách tế bào (callose deposition) và sản sinh các chất oxy hóa (ROS).
- ETS (Effector-Triggered Susceptibility): Để vượt qua PTI, Oomycetes tiết ra các protein độc lực gọi là Effectors (ví dụ: họ protein RxLR ở Phytophthora và sương mai). Các effectors này xâm nhập vào tế bào thực vật, tấn công vào các nút thắt trong hệ thống tín hiệu miễn dịch, làm tê liệt khả năng phòng vệ của cây, cho phép nấm bệnh phát triển.
- ETI (Effector-Triggered Immunity): Nếu cây trồng sở hữu các Gen kháng (R-genes) tương ứng (ví dụ: gen R1, R2 ở khoai tây), các protein kháng này sẽ nhận diện trực tiếp hoặc gián tiếp các Effectors của nấm. Sự nhận diện này kích hoạt phản ứng miễn dịch mạnh mẽ và đặc hiệu gọi là ETI, thường dẫn đến Phản ứng Quá mẫn (Hypersensitive Response - HR) tế bào cây tự chết theo chương trình tại điểm nhiễm bệnh để cô lập và bỏ đói mầm bệnh.
Các chi Oomycetes trọng yếu
Chi Phytophthora
Chi Pythium
Chi Plasmopara
Chi Peronospora
Chi Pseudoperonospora
Chi Bremia
Chi Peronosclerospora
Chi Albugo
Chi Aphanomyces
Quản lý hóa học và chiến lược kiểm soát bền vững
Do sự khác biệt căn bản về sinh học tế bào (vách cellulose, thiếu ergosterol), các thuốc trừ nấm thông thường thường không hiệu quả với Oomycetes. Việc quản lý dựa trên các nhóm thuốc chuyên biệt gọi là Oomycides.
Nhóm Phenylamides - FRAC 4: Metalaxyl, Metalaxyl-M...
Nhóm Quinone inside Inhibitors- FRAC 21: Cyazofamid, Amisulbrom.
Nhóm Cyanoacetamide-oximes - FRAC 27: Cymoxanil.
Nhóm Carbamates - FRAC 28: Propamocarb (Propamocarb hydrochloride).
Nhóm Carboxylic Acid Amides - FRAC 40: Dimethomorph, Mandipropamid, Iprovalicarb, Benthiavalicarb...
Nhóm Oxysterol Binding Protein Inhibitors - FRAC 49: Oxathiapiprolin, Fluoxapiprolin...
Nhóm Phosphonates - FRAC P07: Fosetyl-Aluminium, Phosphorous acid (Potassium phosphite)