
Sự biệt hóa tế bào thực vật
Tính toàn năng, chu kỳ tế bào, tín hiệu, điều hòa biểu hiện gen, hình thành mô
Tính toàn năng
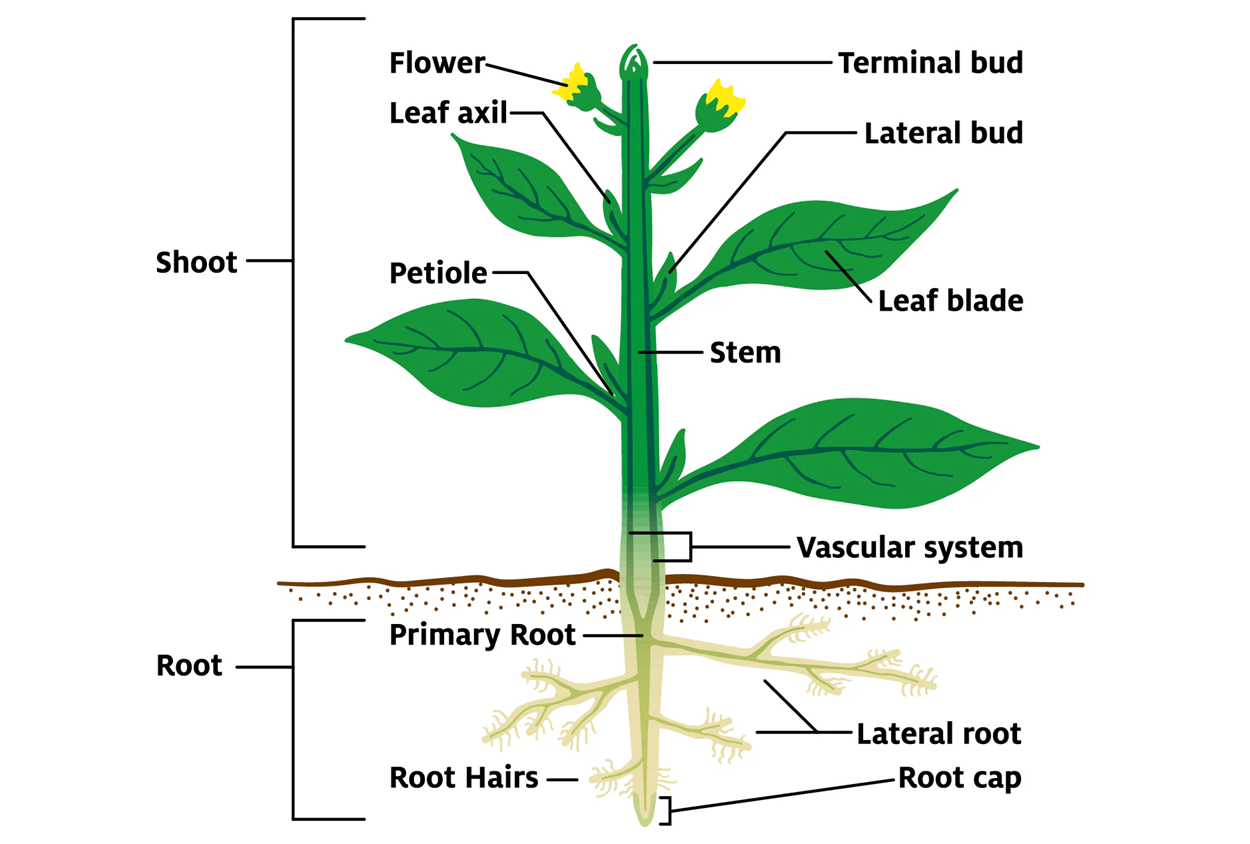
Tính toàn năng của tế bào thực vật (totipotency) là khả năng của một tế bào riêng lẻ, chẳng hạn như tế bào lá, rễ, hoặc thân, khi được đặt trong môi trường dinh dưỡng thích hợp, có thể phân chia, biệt hóa và phát triển thành một cây hoàn chỉnh với đầy đủ các cơ quan (rễ, thân, lá). Đây là đặc điểm cơ bản phân biệt sinh trưởng thực vật với động vật.
Khả năng này tồn tại được là nhờ mỗi tế bào (ngay cả tế bào chuyên hóa như tế bào lá) đều chứa toàn bộ bộ gen cần thiết để mã hóa cho tất cả các bộ phận của cây. Khi gặp điều kiện thích hợp, tế bào đã biệt hóa có thể quên chức năng cũ, quay trở lại trạng thái ban đầu (như tế bào phôi) và bắt đầu phân chia tạo ra mô sẹo (callus). Từ mô sẹo, bằng cách điều chỉnh nồng độ hormone, các tế bào này có thể được lập trình lại để phát triển thành chồi, rễ, và cuối cùng là cây con hoàn chỉnh.
Trong khi tính toàn năng là tiềm năng lý thuyết, thì mô phân sinh (meristems) là biểu hiện vật lý của tiềm năng đó in vivo. Mô phân sinh là các vùng mô chuyên biệt bao gồm các tế bào chưa biệt hóa (giống như tế bào gốc ở động vật), duy trì khả năng phân chia tế bào liên tục trong suốt đời sống của cây. Đây chính là nguồn gốc của mọi sự tăng trưởng về kích thước (chiều dài, thể tích, bề mặt) của cơ thể thực vật.
Mô phân sinh đỉnh (apical meristem) nằm ở đầu tận cùng của thân (đỉnh chồi) và rễ (đỉnh rễ). Hoạt động của mô phân sinh đỉnh chịu trách nhiệm cho sinh trưởng sơ cấp (primary growth), tức là sự gia tăng về chiều dài của thân và rễ. Đây là nơi các cơ quan mới như lá, hoa và rễ bên được khởi tạo.
Mô phân sinh bên (lateral meristem) còn gọi là mô phân sinh thứ cấp, bao gồm tầng phát sinh vỏ (tầng phát sinh bần) và tầng phát sinh trụ (tượng tầng). Mô phân sinh bên chịu trách nhiệm cho sinh trưởng thứ cấp (secondary growth), làm tăng bề ngang (đường kính) của thân và rễ, phổ biến ở cây hai lá mầm và cây hạt trần. Một điểm đáng chú ý là mô phân sinh thứ cấp thường có nguồn gốc từ các tế bào đã phân hóa (như mô mềm) trải qua quá trình "phản biệt hóa" để lấy lại khả năng phân chia, một lần nữa khẳng định nguyên lý tính toàn năng.
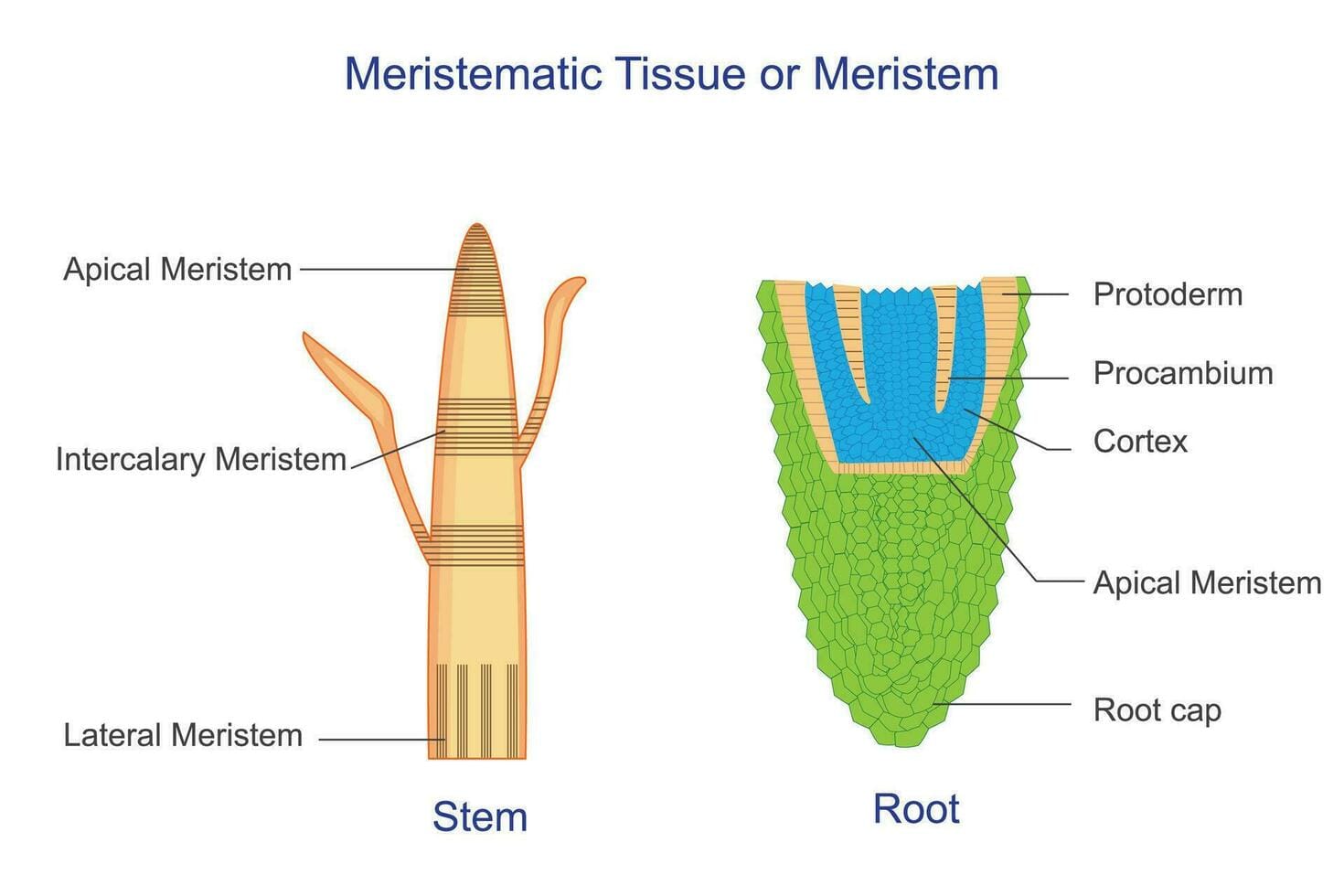
Mô phân sinh lóng (intercalary meristem) đặc trưng cho các cây thuộc họ Một lá mầm, rõ nhất là ở họ Lúa (như lúa, ngô, tre, cỏ). Nằm ở các vị trí xen kẽ, cụ thể là ở gốc của các lóng (khoảng cách giữa hai mấu) trên thân, giúp lóng vươn dài nhanh chóng.
Chu kỳ tế bào
Chu kỳ tế bào là một chuỗi các sự kiện có trật tự nghiêm ngặt, bao gồm hai giai đoạn chính: giai đoạn trung gian (interphase) và giai đoạn phân chia tế bào (M phase, hay nguyên phân).
Kỳ trung gian chiếm phần lớn thời gian trong chu kỳ (thường là hơn 90%). Tế bào không phân chia nhưng hoạt động trao đổi chất rất mạnh mẽ. Kỳ trung gian được chia làm 3 pha nhỏ:
- Pha G1 (Gap 1) là giai đoạn sinh trưởng chính. Tế bào tăng mạnh về kích thước, tổng hợp các protein và bào quan (như ribosome, ty thể...) cần thiết cho sự sống và cho việc nhân đôi ADN sau này.
- Pha S (Synthesis) là sự kiện quan trọng nhất của kỳ trung gian. DNA (vật chất di truyền) được nhân đôi. Kết quả là mỗi nhiễm sắc thể đơn tự nhân đôi thành một nhiễm sắc thể kép, gồm hai chromatid (nhiễm sắc tử chị em) dính nhau ở tâm động.
- Pha G2 (Gap 2), tế bào tiếp tục sinh trưởng nhẹ, tổng hợp các protein cần thiết cho quá trình phân chia (như protein cấu tạo thoi phân bào). Tế bào rà soát lại lần cuối để đảm bảo mọi thứ đã sẵn sàng cho Pha M.
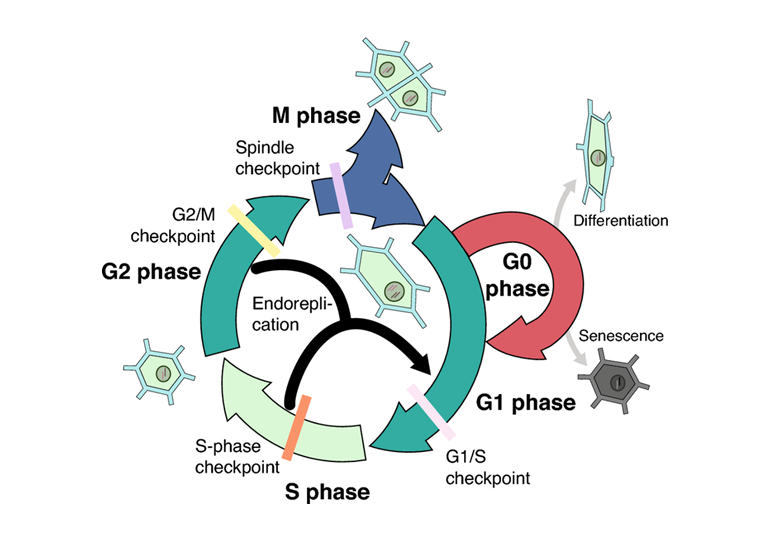
Giai đoạn M, tuy ngắn về thời gian, nhưng lại bao gồm các sự kiện hình thái kịch tính nhất, được chia thành hai quá trình gối đầu lên nhau là phân chia nhân (mitosis) và phân chia tế bào chất (cytokinesis).
- Nguyên phân là quá trình chia nhân, đảm bảo mỗi tế bào con nhận được một bộ nhiễm sắc thể đầy đủ và giống hệt tế bào mẹ. Quá trình này được chia thành 4 kỳ nhỏ (Kỳ đầu, Kỳ giữa, Kỳ sau, Kỳ cuối) để sắp xếp và tách các nhiễm sắc thể kép thành các nhiễm sắc thể đơn rồi di chuyển về hai cực của tế bào.
- Phân chia tế bào chất là quá trình phân chia phần còn lại của tế bào (bào tương và các bào quan).
Sự phân chia tế bào chất ở thực vật có một khác biệt cơ bản so với tế bào động vật, xuất phát từ một ràng buộc vật lý cơ bản: sự tồn tại của thành tế bào (cell wall) cứng nhắc. Do có thành tế bào, tế bào thực vật không thể phân chia bằng cách hình thành eo thắt (cleavage furrow) để co rút màng tế bào lại như ở tế bào động vật. Thay vào đó, tế bào thực vật phải xây dựng một bức tường mới từ bên trong. Cơ chế này được điều khiển bởi một cấu trúc vi ống (microtubule) phức tạp và độc đáo gọi là thể dải phân (phragmoplast). Sự định hướng của mặt phẳng phân chia (được quyết định bởi vị trí hình thành phragmoplast) là một sự kiện hình thái học (morphogenetic) then chốt. Sự phân chia đối xứng hay không đối xứng, và theo mặt phẳng nào (ngang, dọc, hay tiếp tuyến) sẽ quyết định trực tiếp đến hình dạng của mô và cơ quan (ví dụ: tạo ra một chuỗi tế bào, một lớp biểu bì, hay một khối mô đặc).
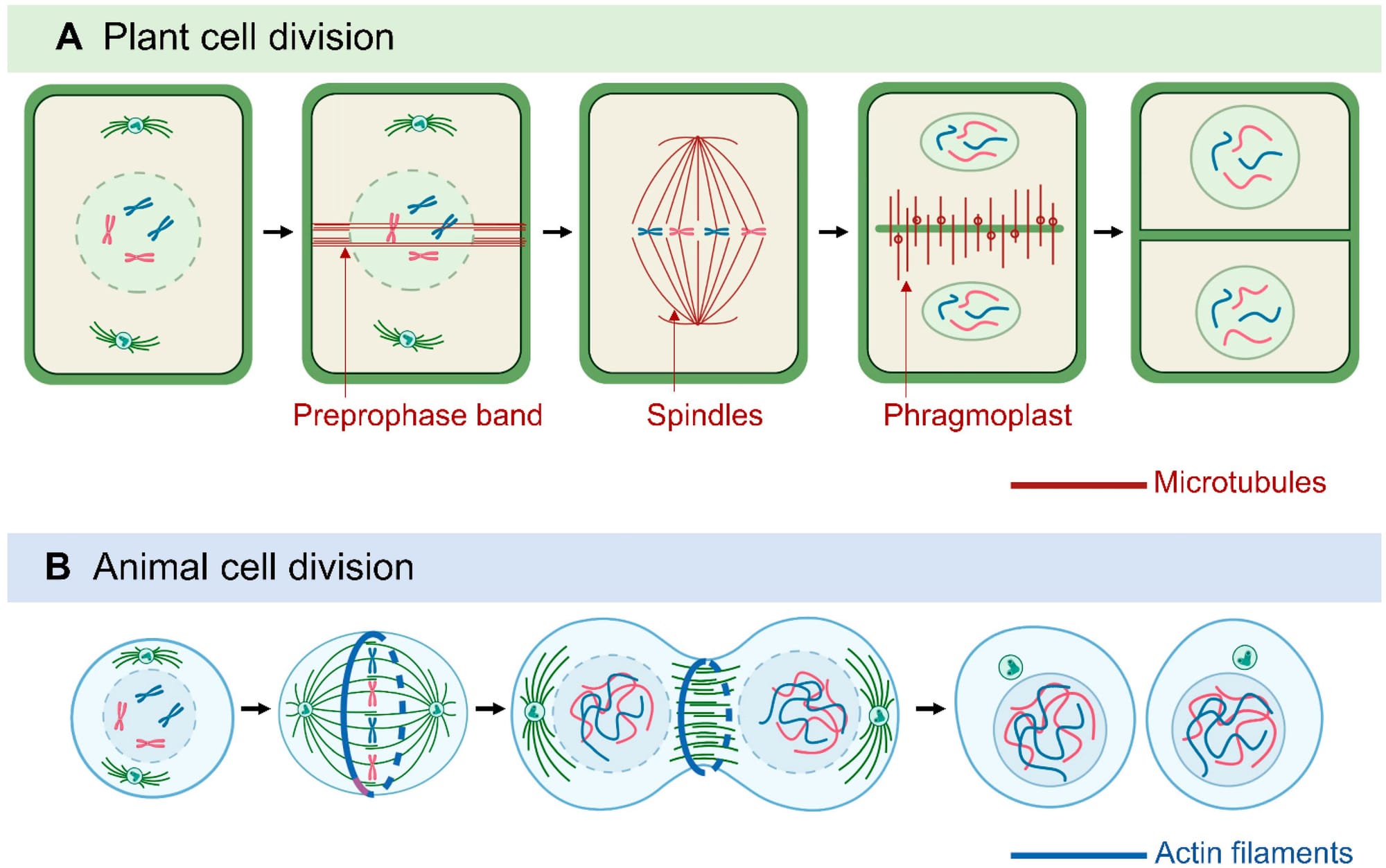
Chu kỳ tế bào không phải là một quá trình tự động. Nó được điều hòa chặt chẽ bởi các tín hiệu bên trong và bên ngoài tế bào. Các điểm kiểm soát tồn tại trong chu kỳ để đảm bảo rằng các sự kiện (như nhân đôi DNA, sắp xếp NST) đã hoàn tất và chính xác trước khi chuyển sang giai đoạn tiếp theo, giúp duy trì tính ổn định của bộ gen.
Sự sinh trưởng của cơ thể thực vật là kết quả của hai quá trình phối hợp chặt chẽ: (1) tăng số lượng tế bào thông qua nguyên phân, và (2) tăng kích thước của các tế bào đó thông qua sự kéo dài.
Nguyên phân (phân bào nguyên nhiễm) tạo ra vật liệu là các tế bào mới. Tuy nhiên, nếu chỉ có phân chia, thực vật sẽ chỉ là một khối tế bào dày đặc. Sự tăng trưởng thực sự về kích thước và việc hình thành các cấu trúc như thân, rễ, lá chủ yếu là do sự kéo dài của các tế bào đó. Hai quá trình này được điều hòa bởi các phytohormone khác nhau:
- Auxin là yếu tố chính thúc đẩy sự kéo dài tế bào.
- Cytokinin là yếu tố chính thúc đẩy sự phân chia tế bào.
Sự phối hợp giữa hai hoạt động này tạo nên nền tảng cho sự phát triển hình thái (morphogenesis) của thực vật.
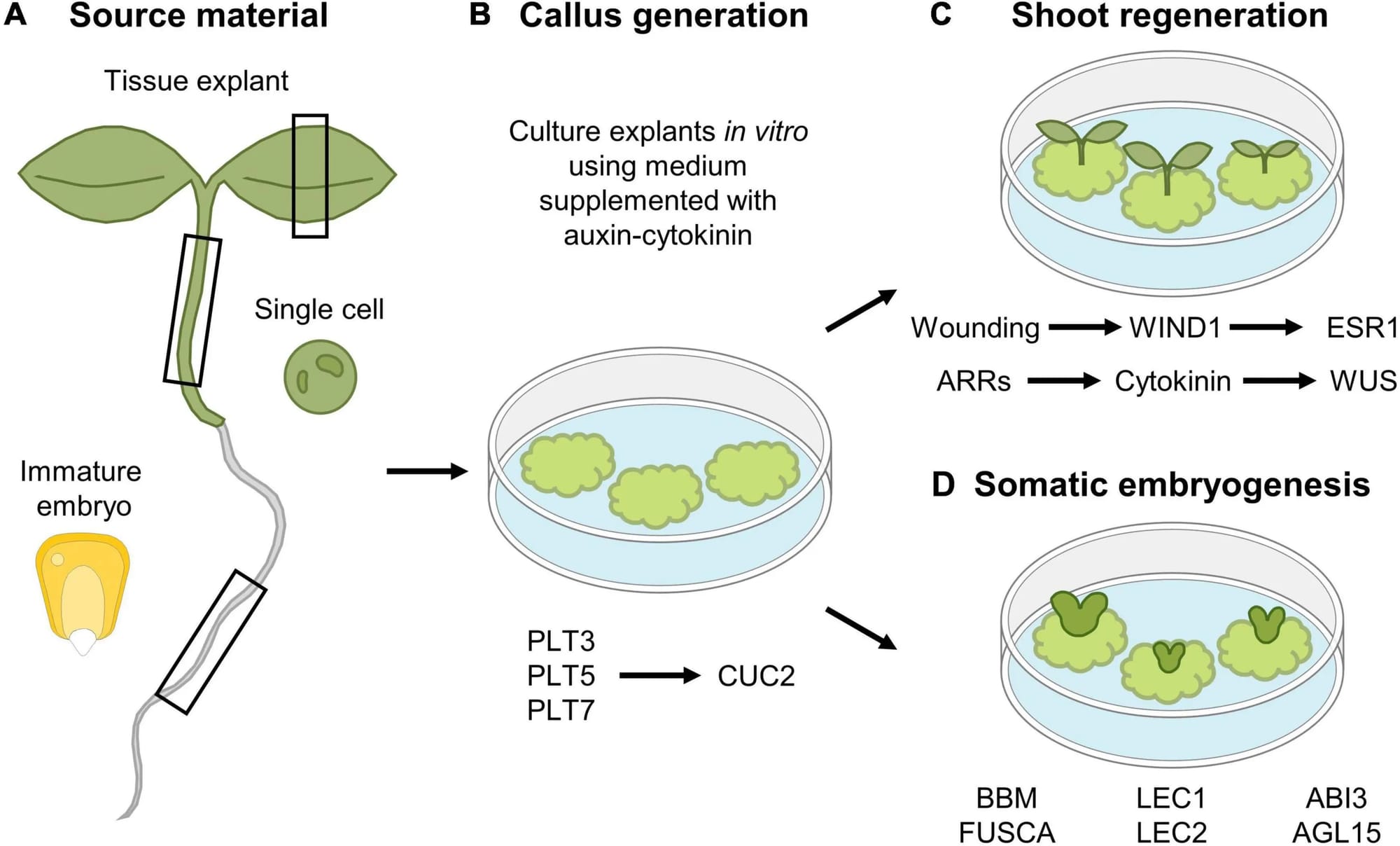
Thông thường, khi một tế bào đã biệt hóa cao, nó sẽ thoát ra khỏi chu kỳ tế bào và ngừng phân chia. Nó đi vào một trạng thái gọi là pha G0 (G-zero). Tuy nhiên, Khi cây bị thương, các tế bào mô mềm (parenchyma) còn sống ở gần vết thương sẽ được kích hoạt. Chúng thoát khỏi G0, quay lại chu kỳ tế bào, phân chia nhanh chóng để tạo ra mô sẹo (callus) làm lành vết thương. Và đây là cơ sở ứng dụng trong nuôi cây mô bằng cách điều chỉnh môi trường có hormone (Auxin, Cytokinin).
Tín hiệu hóa học và môi trường
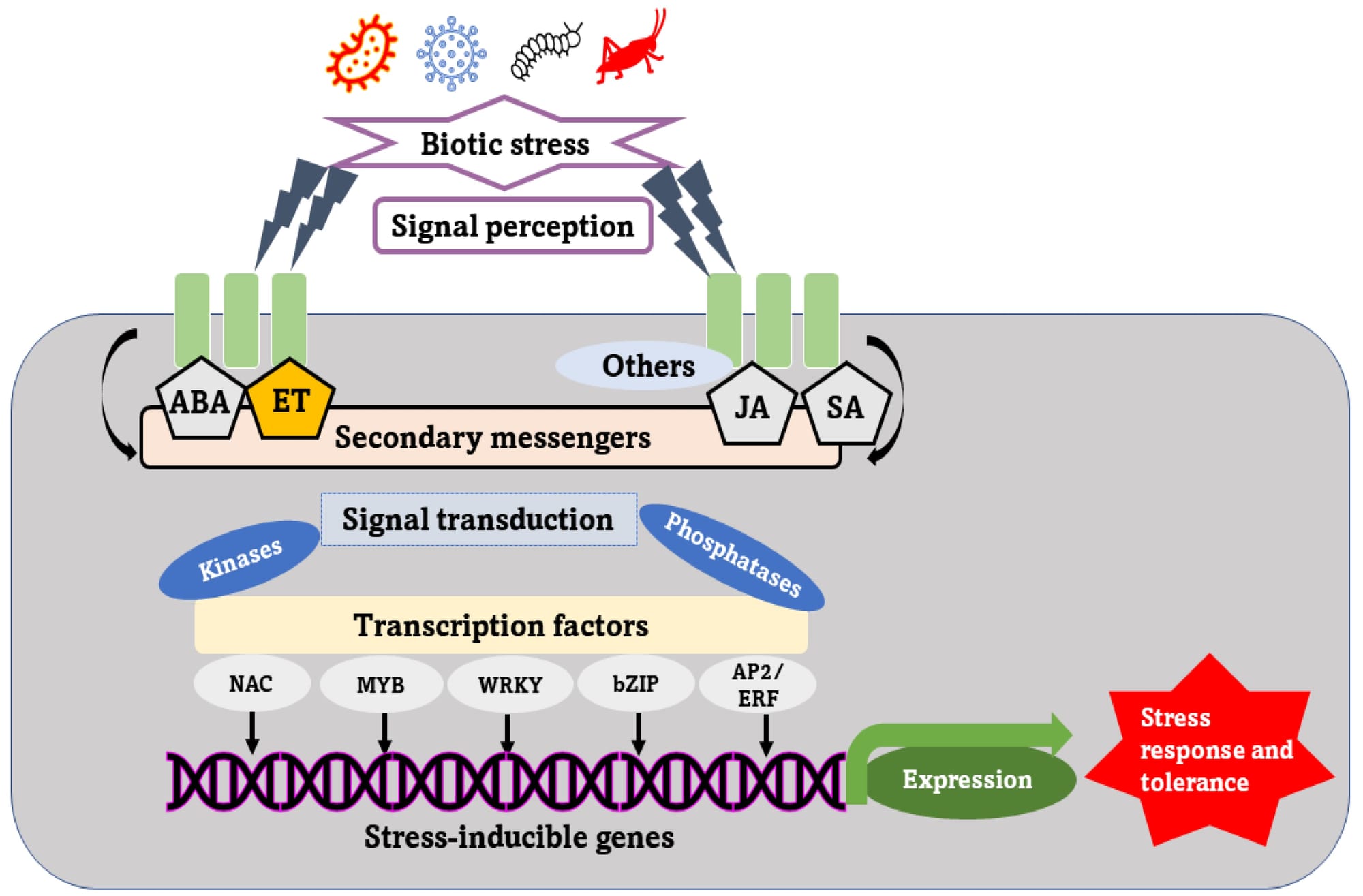
Sự biệt hóa tế bào thực vật (quá trình tế bào đảm nhận một chức năng chuyên biệt, ví dụ như trở thành tế bào rễ, lá, hay mạch dẫn) được điều khiển bởi một mạng lưới phức tạp gồm hai yếu tố chính: tín hiệu hóa học nội sinh (hormone) và tín hiệu môi trường ngoại sinh. Hai yếu tố này không hoạt động độc lập mà tương tác liên tục, cho phép cây thích nghi và phát triển phù hợp với điều kiện xung quanh.
Một trong những phát hiện quan trọng nhất trong sinh lý học thực vật là số phận của một tế bào (ra rễ hay ra chồi) không phụ thuộc vào nồng độ tuyệt đối của một hormone, mà phụ thuộc vào tỷ lệ tương đối giữa Auxin và Cytokinin (tỷ lệ A/C).
Nguyên lý này là nền tảng của công nghệ nuôi cấy mô thực vật (in vitro) :
- Tỷ lệ Auxin cao / Cytokinin thấp (A/C > 1): Tín hiệu này kích thích các tế bào (thường là từ mô sẹo) phân hóa để hình thành rễ.
- Tỷ lệ Cytokinin cao / Auxin thấp (A/C < 1): Tín hiệu này kích thích các tế bào phân hóa để hình thành chồi.
- Tỷ lệ Auxin và Cytokinin cân bằng (A/C ≈ 1): Tỷ lệ này thúc đẩy sự phân chia tế bào nhanh chóng nhưng không phân hóa, tạo ra một khối tế bào không có tổ chức gọi là mô sẹo (callus).
Các hormone quan trọng khác như Gibberellin (GA), Abscisic Acid (ABA), Ethylene cũng hỗ trợ cho sự vươn dài của tế bào, kích thích nảy mầm hoặc biệt hóa để phản ứng với stress và sự chín/già hóa.
Thực vật là sinh vật cố định, không thể di chuyển để trốn tránh điều kiện bất lợi hoặc tìm kiếm môi trường tốt hơn. Do đó, chúng phải điều chỉnh chương trình phát triển (phân chia và phân hóa) để thích ứng với môi trường xung quanh. Ánh sáng là một trong những tín hiệu môi trường quan trọng nhất.
Quá trình phát triển hình thái theo ánh sáng (photomorphogenesis) là một ví dụ điển hình về sự tích hợp tín hiệu. Thực vật sử dụng các protein thụ cảm quang (photoreceptors) chuyên biệt để nhìn thấy ánh sáng. Một trong những nhóm thụ cảm quang quan trọng nhất là Phytochromes, nhạy cảm với ánh sáng đỏ và đỏ xa.
Cơ chế truyền tín hiệu này cho thấy một điểm hội tụ quan trọng: Phytochrome hấp thụ ánh sáng. Khi cảm nhận được ánh sáng, Phytochrome thay đổi cấu hình và di chuyển vào nhân. Tại đây, Phytochrome tương tác và điều chỉnh số lượng hoặc sự ổn định của một nhóm protein khác gọi là các yếu tố phiên mã (Transcription Factors - TFs). Các TFs này sau đó sẽ bật hoặc tắt các gen liên quan đến sự phát triển (ví dụ: gen kiểm soát sự vươn dài của thân hoặc sự mở lá mầm).
Ánh sáng, nhiệt độ, độ ẩm và dinh dưỡng khoáng là những yếu tố môi trường kích hoạt hoặc thay đổi chương trình biệt hóa của cây, thường là thông qua việc thay đổi sản xuất hoặc phân bố hormone.
Điều này cho thấy một nguyên lý điều khiển trung tâm: cả tín hiệu nội sinh (như tỷ lệ A/C) và tín hiệu ngoại sinh (như ánh sáng) đều hội tụ tại một bộ xử lý trung tâm của tế bào. Bộ xử lý đó chính là các yếu tố phiên mã (TFs).
Điều hòa biểu hiện gen
Sự phân hóa về hình thái và chức năng của tế bào là kết quả trực tiếp của sự thay đổi ở cấp độ phân tử. Cốt lõi của sự phân hóa là việc bật và tắt các gen cụ thể. Tất cả các tế bào trong một cơ thể thực vật, từ tế bào lông hút ở rễ đến tế bào mô giậu ở lá, đều chứa cùng một bộ gen (DNA). Sự khác biệt giữa chúng không nằm ở sự hiện diện của gen, mà nằm ở sự biểu hiện của gen.
Ví dụ, một tế bào lá có màu xanh vì nó biểu hiện (bật) các gen cần thiết để tổng hợp lục lạp (chloroplasts) và các enzyme quang hợp. Trong khi đó, một tế bào rễ, mặc dù cũng có các gen đó, nhưng chúng bị tắt (im lặng). Ngược lại, tế bào rễ lại bật các gen chuyên biệt cho việc hấp thụ nước và khoáng.
Quá trình điều hòa này diễn ra ở nhiều cấp độ (sau phiên mã, dịch mã, sau dịch mã), nhưng cấp độ quan trọng nhất và mang tính quyết định nhất là phiên mã (transcription) - quá trình sao chép thông tin từ DNA sang RNA. Kiểm soát khi nào và ở mức độ nào một gen được phiên mã là cách chính mà tế bào thực vật quyết định số phận của nó.
Các yếu tố phiên mã (TFs) là công tắc vật lý thực hiện việc kiểm soát phiên mã. Chúng là các protein có khả năng liên kết với các trình tự DNA đặc hiệu (như promoter hoặc enhancer) nằm gần các gen mục tiêu. Bằng cách liên kết, TFs có thể kích hoạt (tuyển dụng RNA polymerase để bắt đầu phiên mã) hoặc ức chế (ngăn chặn RNA polymerase) sự biểu hiện của gen đó.
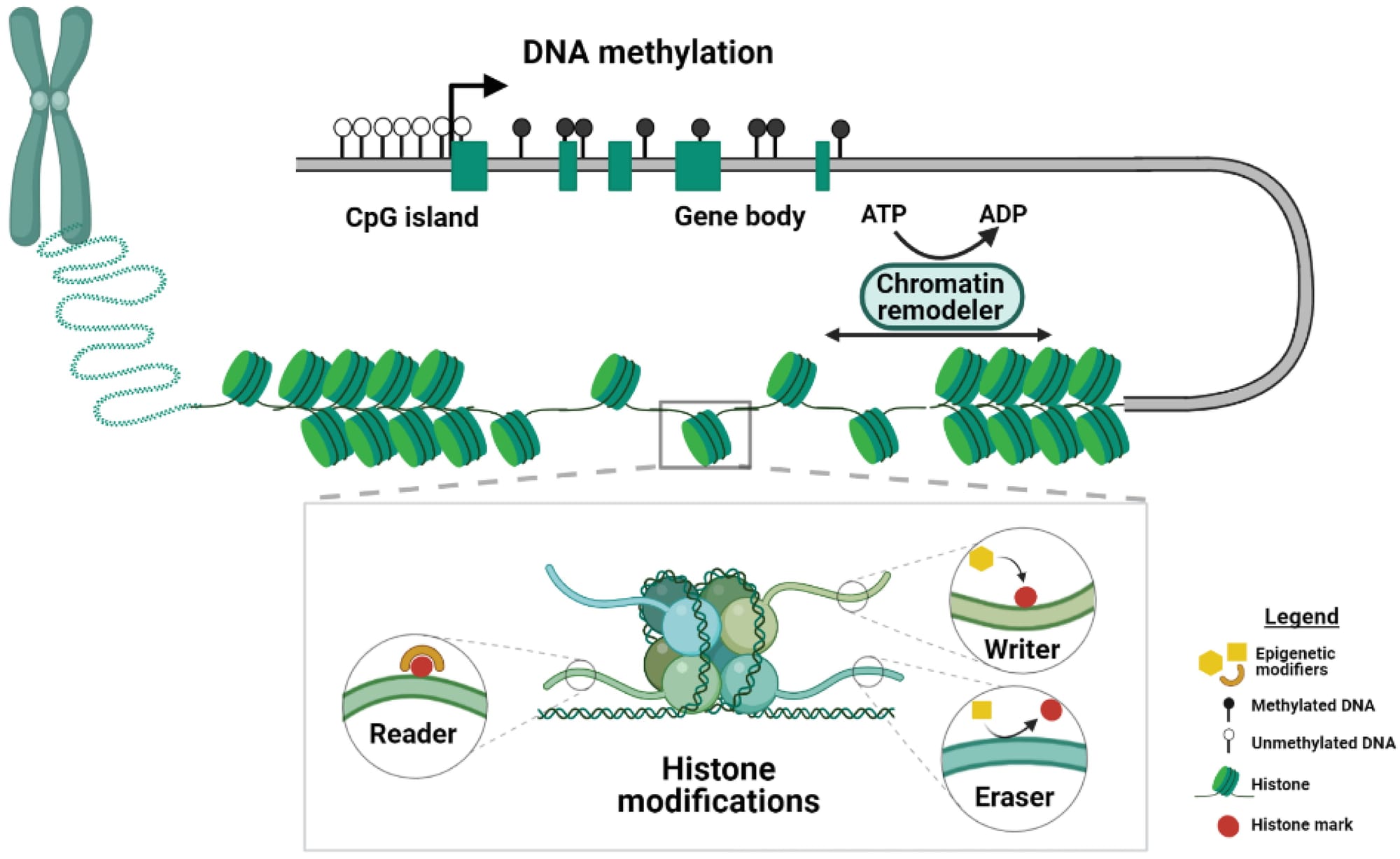
Nếu các TFs là công tắc, thì cơ chế Epigenetic là ổ khóa. Epigenetics đề cập đến các thay đổi di truyền được trong biểu hiện gen mà không làm thay đổi trình tự DNA. Đây là cơ chế bộ nhớ tế bào, đảm bảo rằng một khi tế bào lá đã được quyết định là tế bào lá, nó sẽ duy trì trạng thái đó và truyền trạng thái đó cho các tế bào con của nó qua các lần nguyên phân.
Hai cơ chế epigenetic chính bao gồm:
- Methyl hóa DNA (DNA methylation): Việc gắn một nhóm methyl (CH3) vào các base cytosine trong DNA, đặc biệt là ở các vùng promoter, thường có tác dụng ức chế (làm im lặng) phiên mã gen.
- Tái cấu trúc chromatin (chromatin remodeling) & biến đổi Histone: DNA trong nhân được quấn quanh các protein histone (S18). Cấu trúc DNA-histone này gọi là chromatin. Các phức hợp protein (như DDM1, SWR1) có thể nới lỏng chromatin (euchromatin) để cho phép TFs và RNA polymerase truy cập và phiên mã, hoặc nén chặt chromatin (heterochromatin) để ức chế phiên mã. Khả năng tiếp cận vật lý của TFs với DNA được kiểm soát chặt chẽ bởi trạng thái chromatin.
Sau khi gen đã được phiên mã thành một bản tin (mARN - RNA thông tin), tế bào vẫn có thể kiểm soát nó. Các phân tử RNA nhỏ (như microRNA) có thể tìm và bắt các phân tử mARN cụ thể, có thể phá hủy mARN đó hoặc ngăn không cho nó được dịch mã. Ngay cả khi protein đã được tạo ra, tế bào vẫn kiểm soát hoạt động của nó bằng cách gắn hoặc gỡ bỏ các nhóm hóa học (ví dụ: phosphoryl hóa). Khi protein không còn cần thiết hoặc bị hỏng, tế bào sẽ đánh dấu và phá hủy nó.
Hình thành mô chuyên biệt
Mọi thứ bắt đầu từ mô phân sinh (ví dụ: ở đỉnh chồi, đỉnh rễ). Chúng có khả năng phân chia liên tục và chưa có chức năng cụ thể. Khi nhận được tín hiệu (ví dụ, một phân tử Auxin gắn vào thụ thể), một chuỗi phản ứng được kích hoạt, dẫn đến các yếu tố phiên mã (công tắc gen) đặc hiệu được bật. Các yếu tố này đi vào nhân, tìm và bật một nhóm gen cụ thể trong khi tắt các gen khác.
Các gen được bật sẽ được phiên mã và dịch mã để tạo ra các protein chuyên biệt. Chính các protein này sẽ thay đổi tế bào, thay đổi hình thái, thay đổi chức năng. Khi một nhóm tế bào cùng trải qua quá trình biệt hóa giống nhau, chúng sẽ hình thành một mô chuyên biệt với một chức năng thống nhất. Ví dụ: các tế bào lông hút tạo thành biểu bì rễ (chức năng hút nước), các tế bào ống rỗng, cứng tạo thành mô mạch gỗ (chức năng vận chuyển nước), các tế bào chứa lục lạp tạo thành mô giậu (chức năng quang hợp).

Sự hiểu biết chi tiết về các quá trình này không chỉ mang ý nghĩa học thuật mà còn mở ra những ứng dụng thực tiễn to lớn trong nuôi cấy mô, can thiệp hormone ngoại sinh (PGR) trong canh tác, và kỹ thuật canh tác khoa học hơn.


