
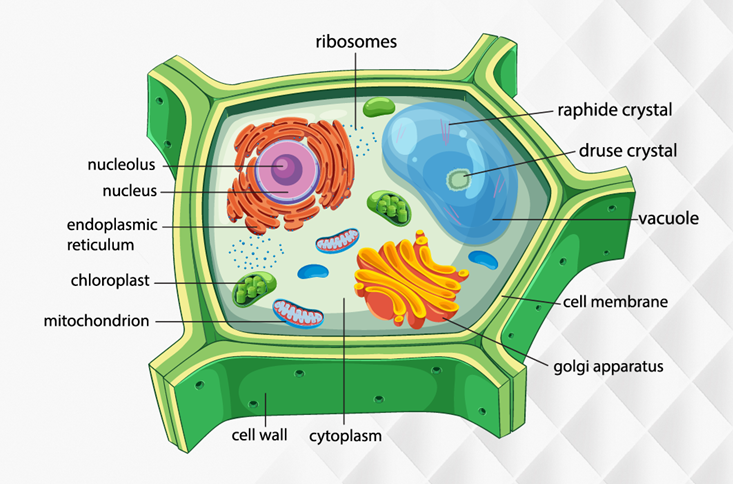
Cấu tạo tế bào thực vật
Thành - Màng - Nhân - Chất.
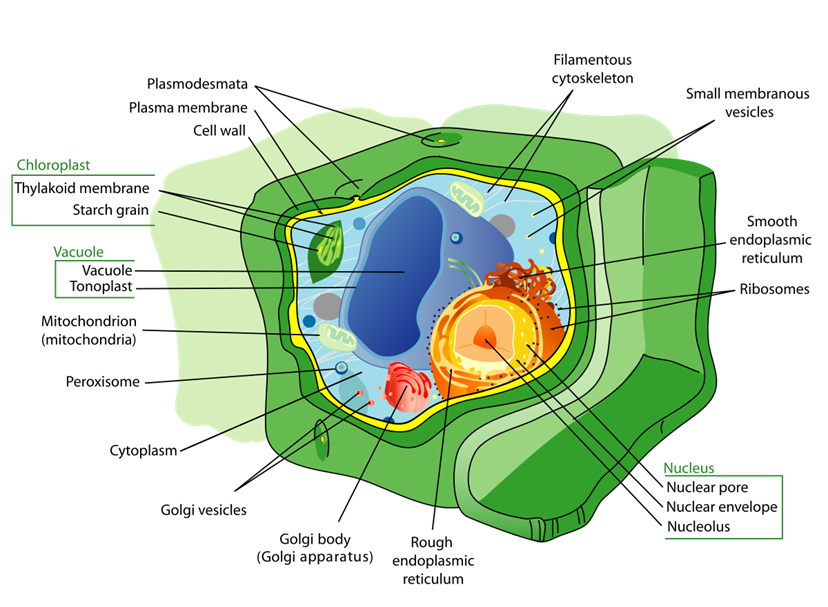
Từ cấu trúc vi mô quyết định chức năng vĩ mô, đến các phản ứng sinh hóa phức tạp điều khiển sự sinh trưởng, ba lĩnh vực: Cấu tạo tế bào, Sinh lý thực vật và Dinh dưỡng là những trụ cột không thể tách rời trong khoa học cây trồng. Hiểu rõ mối quan hệ tương tác giữa chúng là chìa khóa để giải quyết các thách thức trong nông nghiệp và công nghệ sinh học.
Thành tế bào (Cell wall)
Thành tế bào là một trong những đặc điểm nổi bật và quan trọng nhất, phân biệt tế bào thực vật với tế bào động vật. Nó không chỉ là một bức tường cứng bao quanh màng sinh chất mà còn là một cấu trúc siêu vi phức tạp, có tổ chức cao, quyết định đến hình thái và sinh lý của toàn bộ cơ thể thực vật.
Vai trò cơ bản nhất của thành tế bào là cung cấp sự hỗ trợ cơ học và bảo vệ. Nó quy định hình dạng cố định cho tế bào, thường là hình khối hoặc hình đa giác, và bảo vệ các thành phần nguyên sinh chất mỏng manh bên trong khỏi các tác động vật lý và áp suất thẩm thấu. Áp suất trương nước do không bào tạo ra sẽ đẩy vào vách tế bào, tạo ra một độ cứng chắc cho các mô mềm. Chính cấu trúc vững chãi này cho phép thực vật có thể vươn cao và duy trì một cấu trúc thẳng đứng mà không cần đến một hệ thống xương nâng đỡ như ở động vật.
Tuy nhiên, chức năng của thành tế bào vượt xa vai trò của một bộ khung xương ngoại bào. Nó là một cấu trúc năng động, tham gia vào nhiều quá trình sinh lý thiết yếu của thực vật, bao gồm việc điều tiết sự hấp thu nước và khoáng chất, kiểm soát quá trình thoát hơi nước, vận chuyển các chất giữa các tế bào, và tiết ra các hợp chất hóa học. Hơn nữa, thành tế bào còn là hàng rào phòng thủ đầu tiên, giúp bảo vệ cây khỏi sự tấn công của virus và các tác nhân gây bệnh khác. Sự phát triển và biến đổi của thành tế bào cũng đóng vai trò quan trọng trong việc điều chỉnh vòng đời của sinh vật thực vật.
Thành tế bào thực vật là một vật liệu composite sinh học, được cấu thành từ nhiều loại polymer khác nhau, mỗi loại đóng một vai trò chuyên biệt. Thành phần cấu trúc chính, tạo nên bộ khung sườn là cellulose. Cellulose là một polysaccharide được tạo thành từ các chuỗi dài các phân tử β-D-glucose liên kết với nhau. Các chuỗi cellulose này liên kết song song với nhau thông qua các liên kết hydro để tạo thành các cấu trúc siêu bền gọi là vi sợi (microfibrils). Các vi sợi này có độ bền kéo cực cao, tương đương với thép, tạo nên một mạng lưới dai và chắc chắn cho toàn bộ thành tế bào.
Lấp đầy các khoảng trống giữa các vi sợi cellulose là một cơ chất vô định hình, giống như gel, được gọi là matrix. Cơ chất này chủ yếu bao gồm hai loại polysaccharide khác:
- Hemicellulose: Là một nhóm đa dạng các polysaccharide phân nhánh (bao gồm galactose, manose, xylose, arabinose) có khả năng liên kết chéo với các vi sợi cellulose, giúp cố định chúng lại với nhau và tạo thành một mạng lưới ba chiều vững chắc.
- Pectin: Là một nhóm các polysaccharide phức tạp khác, có đặc tính ưa nước và tạo gel cao. Pectin tạo thành một ma trận hydrat hóa, quyết định độ xốp của vách tế bào, qua đó ảnh hưởng đến sự lưu thông của nước và các chất hòa tan. Nó cũng đóng vai trò như một chất kết dính giữa các tế bào.
Tùy thuộc vào loại tế bào và giai đoạn phát triển, vách tế bào có thể được gia cố thêm bởi các hợp chất hóa học khác, một quá trình được gọi là sự tẩm.
- Lignin: Là một polymer thơm phức tạp, có tính kị nước và rất cứng. Lignin được tẩm vào ma trận polysaccharide của vách thứ cấp, đặc biệt là ở các tế bào mô gỗ, làm cho vách trở nên cứng rắn hơn, tăng cường khả năng chống chịu áp lực cơ học và làm cho vách không thấm nước. Quá trình này được gọi là "sự hóa gỗ".
- Suberin và Cutin: Là những chất béo, có tính kị nước cao. Suberin được tẩm vào vách của các tế bào bần (mô bảo vệ ở thân và rễ cây già), trong khi cutin tạo thành một lớp (lớp cutin) phủ bên ngoài vách của các tế bào biểu bì lá. Cả hai chất này đều có chức năng ngăn chặn sự mất nước và bảo vệ cây khỏi các tác nhân gây bệnh từ môi trường.
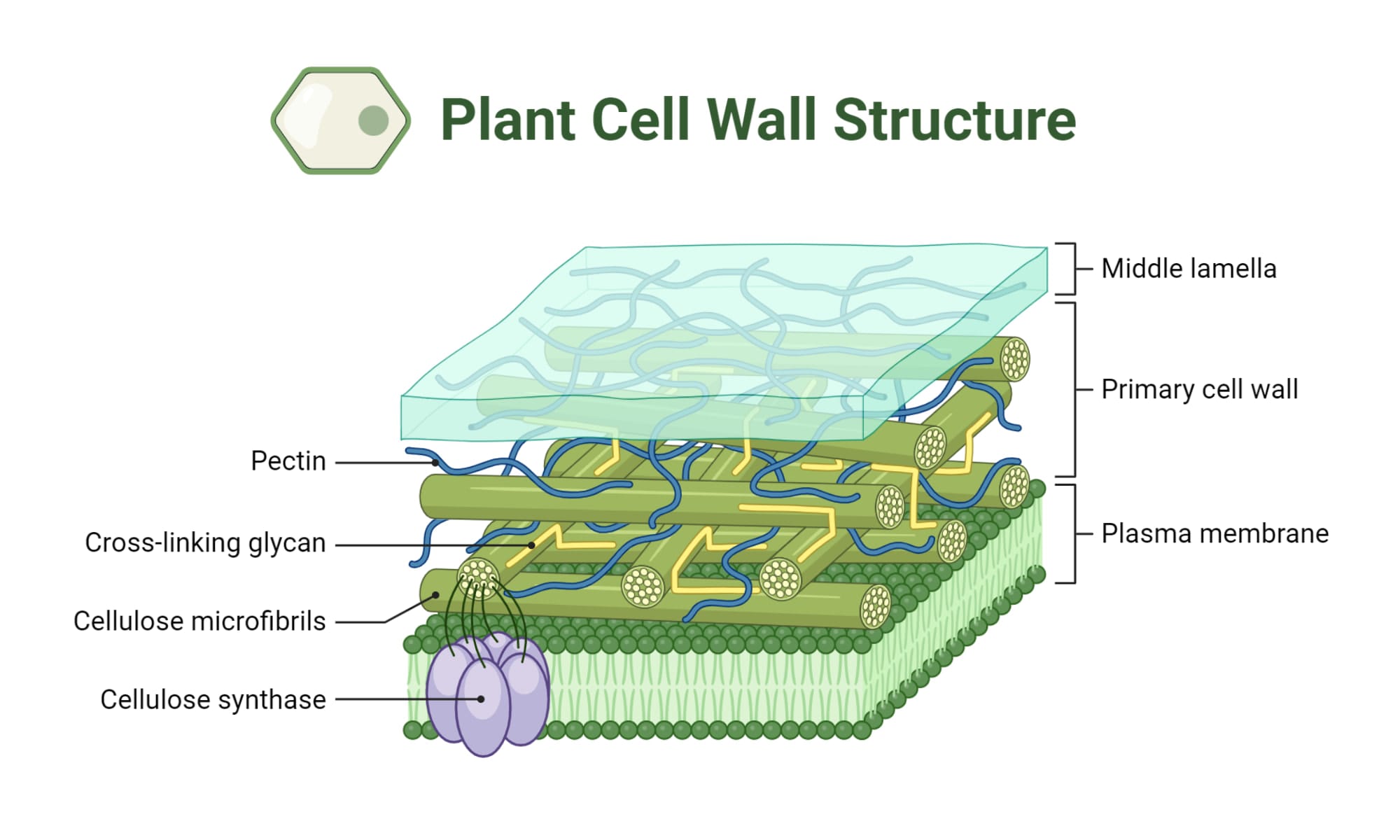
Thành tế bào không phải là một cấu trúc đồng nhất mà được hình thành theo từng lớp trong quá trình phát triển của tế bào.
- Phiến giữa (Middle Lamella): Đây là lớp ngoài cùng của thành tế bào, hình thành đầu tiên trong giai đoạn cuối của quá trình phân bào, có chức năng như một lớp "xi măng" gắn kết các tế bào con lại với nhau. Thành phần chủ yếu của phiến giữa là các hợp chất pectic, có thể kết hợp với các ion canxi để tạo thành canxi pectate, tăng cường độ kết dính.
- Vách sơ cấp (Primary Wall): Được hình thành ngay sau phiến giữa, khi tế bào còn đang trong giai đoạn tăng trưởng và phát triển. Vách sơ cấp có mặt ở tất cả các loại tế bào thực vật. Nó tương đối mỏng, linh hoạt và có khả năng giãn nở để cho phép tế bào tăng kích thước. Về thành phần, vách sơ cấp chứa một tỷ lệ cellulose tương đối thấp (khoảng 8-25%) và một tỷ lệ cao pectin và hemicellulose. Các vi sợi cellulose trong vách sơ cấp thường được sắp xếp một cách ngẫu nhiên, tạo điều kiện cho sự giãn nở đa hướng.
- Vách thứ cấp (Secondary Wall): Sau khi tế bào đã đạt đến kích thước tối đa và ngừng tăng trưởng, nó có thể bắt đầu hình thành vách thứ cấp. Lớp vách này được bồi đắp vào phía bên trong của vách sơ cấp, nằm giữa vách sơ cấp và màng sinh chất. Vách thứ cấp dày và cứng chắc hơn nhiều so với vách sơ cấp. Nó có hàm lượng cellulose cao hơn đáng kể (30-50% hoặc hơn) và hàm lượng pectin thấp hơn. Đặc biệt, vách thứ cấp thường được tẩm thêm lignin, làm tăng cường độ cứng và khả năng chống thấm. Một đặc điểm quan trọng của vách thứ cấp là cấu trúc nhiều lớp. Trong mỗi lớp, các vi sợi cellulose được sắp xếp song song với nhau, nhưng hướng của các vi sợi lại khác nhau giữa các lớp liền kề, thường chéo nhau một góc từ 60 đến 90 độ. Cấu trúc nhiều lớp composite này mang lại độ bền cơ học vượt trội, chống lại lực tác động từ nhiều hướng.
Mặc dù thành tế bào tạo ra một rào cản đáng kể, sự giao tiếp và vận chuyển giữa các tế bào vẫn được duy trì thông qua các cấu trúc chuyên biệt.
- Lỗ (Pits): Quá trình bồi đắp vách thứ cấp thường không đồng đều, để lại những vùng mỏng nơi chỉ có vách sơ cấp và phiến giữa. Những vùng này được gọi là lỗ. Các lỗ của hai tế bào lân cận thường nằm đối diện nhau, tạo thành một cặp lỗ, cho phép sự thông thương dễ dàng hơn giữa chúng.
- Cầu sinh chất (Plasmodesmata): Đây là những kênh giao tiếp trực tiếp, là những ống nhỏ xuyên qua vách tế bào, nối liền tế bào chất của các tế bào lân cận với nhau. Cấu trúc này cho phép sự trao đổi trực tiếp các phân tử nhỏ và thậm chí cả các đại phân tử, đóng vai trò sống còn trong việc phối hợp hoạt động của các tế bào trong một mô. Ngay cả khi tế bào bị co nguyên sinh, tưởng chừng như tế bào chất đã tách rời khỏi vách, sự tiếp xúc vẫn được duy trì thông qua các cầu sinh chất này.
Quan niệm truyền thống thường xem thành tế bào như một cấu trúc chết, một cái hộp trơ ì bao bọc phần sống bên trong. Tuy nhiên, các bằng chứng hiện đại cho thấy một bức tranh phức tạp và năng động hơn nhiều. Quá trình tổng hợp và tái cấu trúc thành tế bào là một quá trình sinh hóa được điều khiển chặt chẽ bởi tế bào, phụ thuộc vào giai đoạn phát triển và các tín hiệu từ môi trường. Sự thay đổi thành phần hóa học, như hóa gỗ hay hóa bần, là những đáp ứng sinh lý tích cực. Hơn nữa, sự tồn tại của các kết nối vật lý và chức năng liên tục với tế bào chất thông qua các cầu sinh chất cho thấy thành tế bào không hề bị cô lập. Do đó, một cách tiếp cận chính xác hơn là xem thành tế bào như một bào quan ngoại bào (extracellular organelle) - một cấu trúc động, có chức năng trao đổi chất, và là một phần mở rộng không thể thiếu của tế bào sống.
Bên cạnh đó, cấu trúc của thành tế bào là một minh chứng tuyệt vời cho nguyên lý kỹ thuật vật liệu composite trong tự nhiên. Sự kết hợp giữa các vi sợi cellulose có độ bền kéo cao (tương tự sợi gia cường) được nhúng trong một ma trận pectin và hemicellulose có tính kết dính và chịu nén (tương tự nhựa nền) tạo ra một vật liệu vừa nhẹ vừa cực kỳ bền chắc. Đặc biệt, cấu trúc nhiều lớp của vách thứ cấp, với các hướng sợi xen kẽ , tương tự như cách con người chế tạo ván ép hay vật liệu composite sợi carbon để tối đa hóa khả năng chống chịu lực từ nhiều hướng. Giải pháp kỹ thuật vật liệu tinh vi này chính là yếu tố cho phép thực vật xây dựng nên những cơ thể khổng lồ, vươn cao hàng chục mét để cạnh tranh ánh sáng, chống lại trọng lực và các tác động của môi trường.
Màng sinh chất (Cell membrane)
Nằm ngay bên trong thành tế bào, bao bọc lấy toàn bộ tế bào chất, là màng sinh chất hay còn gọi là màng plasma. Đây là ranh giới thực sự giữa môi trường nội bào và ngoại bào, một cấu trúc linh hoạt và có tính chọn lọc cao, đóng vai trò then chốt trong việc duy trì sự sống của tế bào.
Nền tảng của màng sinh chất là lớp kép phospholipid. Mỗi phân tử phospholipid có tính lưỡng cực, bao gồm một đầu ưa nước chứa nhóm phosphate và hai đuôi kị nước là các chuỗi hydrocarbon. Trong môi trường nước của tế bào, các phân tử phospholipid tự sắp xếp thành một lớp kép, với các đuôi kị nước quay vào trong, tránh xa nước, và các đầu ưa nước quay ra ngoài, tiếp xúc với môi trường nước ở cả bên trong và bên ngoài tế bào. Cấu trúc này tạo ra một lõi kị nước liên tục ở giữa màng, hoạt động như một rào cản hiệu quả, ngăn cản sự di chuyển tự do của các ion và các phân tử phân cực, tan trong nước.
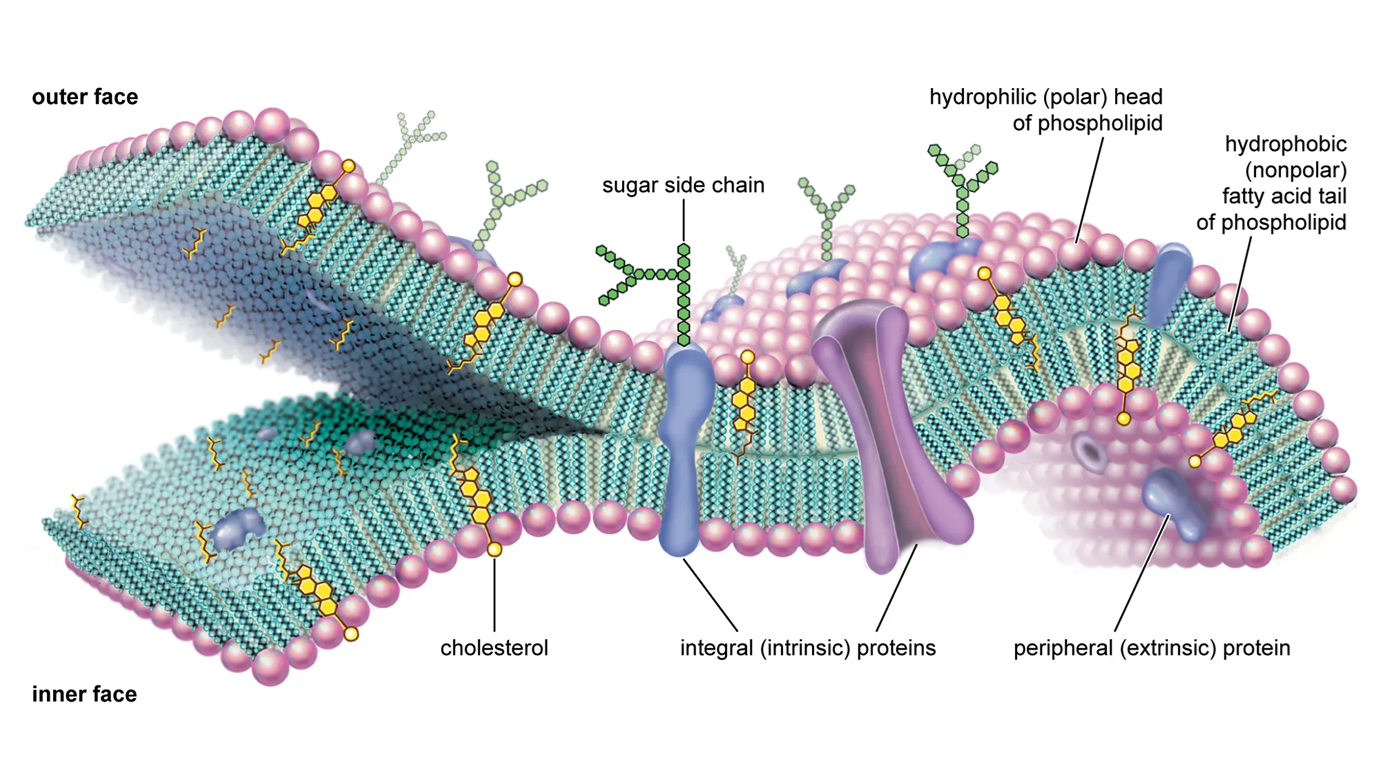
Trong khi lớp kép phospholipid tạo ra cấu trúc cơ bản và tính thấm của màng, thì các protein màng lại thực hiện hầu hết các chức năng chuyên biệt. Chúng đóng vai trò như những "công nhân" của màng, đảm nhiệm các nhiệm vụ đa dạng như:
- Vận chuyển: Tạo ra các kênh hoặc hoạt động như các chất mang để vận chuyển có chọn lọc các ion và phân tử nhỏ qua màng.
- Hoạt tính enzyme: Một số protein màng là các enzyme, xúc tác cho các phản ứng hóa học diễn ra tại bề mặt màng.
- Truyền tín hiệu: Các protein thụ thể trên màng có khả năng liên kết với các phân tử tín hiệu từ bên ngoài (như hormone thực vật) và khởi động một chuỗi các phản ứng truyền tin vào bên trong tế bào.
- Nhận biết tế bào: Các glycoprotein (protein liên kết với carbohydrate) trên bề mặt màng có thể đóng vai trò là "dấu chuẩn" giúp các tế bào nhận biết lẫn nhau.
Cấu trúc của màng sinh chất được mô tả một cách thành công nhất bởi mô hình khảm động, do S. J. Singer và G. L. Nicolson đề xuất vào năm 1972. Mô hình này nhấn mạnh hai đặc tính cơ bản của màng: tính "khảm" và tính "động".
Tính "Khảm": Màng không phải là một cấu trúc đồng nhất mà được cấu tạo từ nhiều loại phân tử khác nhau "khảm" vào nhau. Thành phần chính bao gồm một lớp kép phospholipid và các phân tử protein. Các protein này không được sắp xếp một cách cố định mà phân bố rải rác trong lớp kép lipid, giống như những viên đá khảm trên một bức tranh. Có hai loại protein màng chính: protein xuyên màng, đi qua toàn bộ chiều dày của màng, và protein ngoại vi, chỉ bám vào bề mặt trong hoặc ngoài của màng.
Tính "Động": Các thành phần của màng không ở trạng thái tĩnh mà có khả năng di chuyển tương đối với nhau. Các phân tử phospholipid có thể di chuyển ngang trong lớp của chúng, quay quanh trục và thậm chí (hiếm khi) lật từ lớp này sang lớp kia. Các protein màng cũng có thể trôi nổi tự do trong "biển" lipid. Sự linh hoạt này là do các phân tử trong màng được liên kết với nhau chủ yếu bằng các tương tác kị nước và các liên kết yếu khác, chứ không phải bằng các liên kết cộng hóa trị bền chặt.
Từ cấu trúc khảm-động, hai chức năng cơ bản của màng sinh chất được hình thành:
- Điều hòa vận chuyển: Đây là chức năng quan trọng nhất. Màng sinh chất hoạt động như một rào cản có tính thấm chọn lọc, nghĩa là nó cho phép một số chất đi qua dễ dàng trong khi ngăn chặn hoặc kiểm soát chặt chẽ sự di chuyển của các chất khác. Sự kiểm soát này là tối quan trọng để tế bào có thể duy trì một môi trường nội bào ổn định, có thành phần hóa học khác biệt và tối ưu cho các hoạt động sống, bất chấp sự thay đổi của môi trường bên ngoài.
- Trung tâm giao tiếp: Màng sinh chất là giao diện của tế bào với thế giới bên ngoài. Nó không chỉ tiếp nhận các tín hiệu hóa học mà còn tham gia vào việc truyền các tín hiệu vật lý, tương tác với các tế bào lân cận và với ma trận ngoại bào (trong trường hợp này là thành tế bào).
Một khía cạnh sâu sắc hơn của cấu trúc màng là tính bất đối xứng, vốn là nền tảng cho chức năng định hướng của nó. Hai lớp của màng sinh chất không hoàn toàn giống nhau về thành phần lipid và protein. Ví dụ, các chuỗi carbohydrate của glycoprotein và glycolipid thường chỉ được tìm thấy ở mặt ngoài của màng, tạo thành một lớp gọi là glycocalyx. Các protein thụ thể có các vùng chức năng riêng biệt, một vùng hướng ra ngoài để nhận tín hiệu và một vùng hướng vào trong để tương tác với các phân tử nội bào. Các quá trình vận chuyển tích cực, như bơm ion, cũng có tính định hướng rõ rệt. Sự bất đối xứng này là thiết yếu, nó biến màng từ một rào cản đơn thuần thành một giao diện có "hướng", phân biệt rõ ràng "bên trong" và "bên ngoài", với mặt ngoài chuyên về tiếp nhận thông tin và nhận diện, còn mặt trong chuyên về tương tác với bộ khung tế bào và các con đường truyền tín hiệu nội bào.
Bên cạnh đó, tính "động" hay tính lỏng của màng là một đặc tính vật lý được tối ưu hóa qua quá trình tiến hóa, thể hiện một sự đánh đổi có chủ đích giữa sự ổn định và chức năng. Nếu màng quá cứng, các protein sẽ không thể di chuyển và tương tác với nhau, làm tê liệt nhiều chức năng quan trọng như truyền tín hiệu hay vận chuyển. Ngược lại, nếu màng quá lỏng, nó sẽ mất đi tính toàn vẹn cấu trúc và không thể duy trì vai trò rào cản. Tế bào điều chỉnh tính lỏng của màng bằng cách thay đổi thành phần axít béo trong phospholipid (các axít béo không no làm tăng tính lỏng) và hàm lượng các phân tử sterol (ở tế bào động vật là cholesterol, ở thực vật là các phytosterol khác) xen kẽ vào lớp lipid để tăng cường sự ổn định. Do đó, tính lỏng của màng không phải là một hằng số mà là một đặc tính được điều chỉnh linh hoạt để cân bằng giữa việc duy trì một cấu trúc bền vững và cho phép các hoạt động sinh hóa năng động cần thiết cho sự sống.
Nhân tế bào (Nucleus)
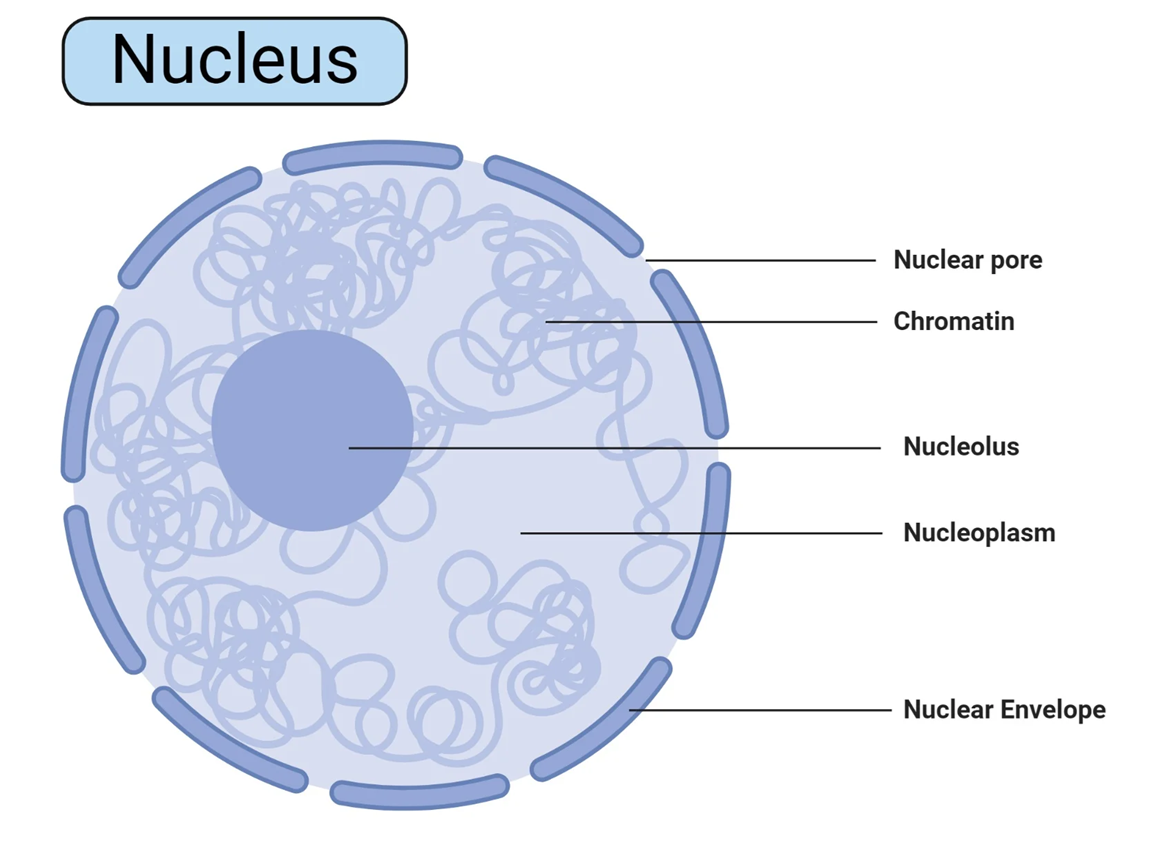
Trong thế giới các bào quan, nhân tế bào là cấu trúc nổi bật và quan trọng nhất, thường được ví như "bộ não" hay "trung tâm chỉ huy" của tế bào. Nhân là bào quan lớn nhất, thường có dạng hình cầu, và là nơi chứa đựng gần như toàn bộ vật chất di truyền của tế bào dưới dạng các phân tử DNA. Vai trò của nhân không chỉ dừng lại ở việc lưu trữ thông tin. Nó còn là trung tâm quản trị, điều hành và giám sát mọi hoạt động sống của tế bào, từ tăng trưởng, trao đổi chất, tổng hợp protein cho đến quá trình phức tạp nhất là phân chia tế bào. Bằng cách kiểm soát sự biểu hiện của các gen, tức là quyết định gen nào được "bật" hay "tắt" tại thời điểm nào → nhân điều khiển việc sản xuất các protein và enzyme, qua đó định hình cấu trúc và chức năng của tế bào.
Bao bọc và bảo vệ vật chất di truyền bên trong là màng nhân, một cấu trúc màng kép độc đáo. Màng nhân bao gồm hai lớp màng lipid kép song song, màng trong và màng ngoài ngăn cách nhau bởi một khoảng không gian gọi là xoang quanh nhân. Mỗi lớp màng có độ dày khoảng 6-9 nm. Màng ngoài của nhân có một đặc điểm cấu trúc quan trọng là nó liên tục với màng của lưới nội chất hạt, cho thấy một mối liên kết vật lý và chức năng chặt chẽ trong hệ thống nội màng của tế bào.
Màng nhân không phải là một lớp vỏ kín hoàn toàn. Nó được đục thủng bởi hàng ngàn cấu trúc cực kỳ phức tạp gọi là phức hợp lỗ nhân. Mỗi lỗ nhân không phải là một cái lỗ trống mà là một cỗ máy phân tử khổng lồ, được cấu tạo từ hàng chục loại protein khác nhau (gọi là nucleoporin). Những phức hợp này hoạt động như những cổng gác có khả năng điều tiết cao độ, kiểm soát chặt chẽ sự vận chuyển hai chiều của các phân tử giữa nhân và tế bào chất. Các phân tử nhỏ và ion có thể khuếch tán tự do qua lỗ, nhưng các đại phân tử như protein (cần cho hoạt động của nhân) và RNA (mang thông tin di truyền ra ngoài) phải được vận chuyển một cách tích cực và có chọn lọc thông qua sự tương tác với các protein của phức hợp lỗ nhân.
Cấu trúc màng kép và hệ thống lỗ nhân tinh vi này phản ánh một giải pháp tiến hóa xuất sắc cho một vấn đề nan giải: làm thế nào để vừa tách biệt, bảo vệ bộ gen quý giá khỏi môi trường hóa học phức tạp của tế bào chất, lại vừa đảm bảo sự giao tiếp liên tục và có kiểm soát giữa hai khu vực này. Màng nhân tạo ra một rào cản vật lý hiệu quả để bảo vệ DNA. Tuy nhiên, tế bào không thể hoạt động nếu nhân bị cô lập hoàn toàn. Thông tin di truyền phải được truyền ra ngoài dưới dạng mRNA, và các enzyme, protein cấu trúc cần thiết cho việc sao chép và duy trì DNA phải được nhập vào trong. Phức hợp lỗ nhân chính là giải pháp cho mâu thuẫn này. Nó không phải là một khiếm khuyết của màng mà là một cơ chế kiểm soát tinh vi, cho phép sự giao tiếp chọn lọc và được điều tiết, cân bằng hoàn hảo giữa việc bảo vệ bộ gen và việc tích hợp chức năng của nhân vào hoạt động chung của tế bào.
Bên trong nhân là chất nhiễm sắc (chromatin), một phức hợp gồm các phân tử DNA mạch kép, dài, liên kết với một lượng lớn các protein, trong đó quan trọng nhất là các protein kiềm tính gọi là histone. Sự gói ghém này có vai trò kép. Thứ nhất, nó giúp nén gọn một cách đáng kinh ngạc các phân tử DNA khổng lồ (có thể dài tới hàng mét nếu duỗi thẳng) vào bên trong một không gian cực nhỏ của nhân (chỉ vài micromet đường kính). Thứ hai, và quan trọng không kém, mức độ gói ghém của chất nhiễm sắc đóng vai trò then chốt trong việc điều hòa hoạt động của gen.
Khi tế bào không phân chia (ở giai đoạn gian kỳ/kỳ trung gian), chất nhiễm sắc tồn tại ở dạng các sợi mảnh, phân tán. Tuy nhiên, khi tế bào chuẩn bị bước vào quá trình phân chia, các sợi chất nhiễm sắc này sẽ trải qua một quá trình xoắn và cô đặc ở nhiều cấp độ, cuối cùng tạo thành các cấu trúc riêng rẽ, có hình dạng đặc trưng gọi là nhiễm sắc thể, có thể quan sát được dưới kính hiển vi quang học. Mỗi loài sinh vật nhân thực có một số lượng và hình thái nhiễm sắc thể đặc trưng.
Việc tổ chức chất nhiễm sắc không chỉ đơn thuần là một giải pháp đóng gói DNA. Nó còn là một cơ chế điều hòa biểu hiện gen ở cấp độ cao. Các vùng chất nhiễm sắc có thể tồn tại ở trạng thái "mở" hơn, ít xoắn hơn, gọi là chất nhiễm sắc thật (euchromatin), cho phép các enzyme phiên mã dễ dàng tiếp cận và "đọc" các gen trong vùng đó. Ngược lại, các vùng được gói ghém chặt chẽ hơn, gọi là chất dị nhiễm sắc (heterochromatin), thường chứa các gen không hoạt động hoặc bị "tắt". Mức độ xoắn và cô đặc của chất nhiễm sắc không phải là cố định mà có thể thay đổi một cách linh hoạt dưới tác động của các yếu tố điều hòa, qua đó kiểm soát việc gen nào được biểu hiện tại một thời điểm nhất định. Do đó, cấu trúc vật lý của chất nhiễm sắc là một lớp thông tin bổ sung (thông tin biểu sinh) quan trọng, ảnh hưởng trực tiếp đến chức năng và số phận của tế bào.
Bên trong nhân, có thể quan sát thấy một hoặc một vài cấu trúc hình cầu, bắt màu đậm hơn so với phần còn lại, đó là nhân con. Nhân con không có màng bao bọc và được cấu tạo chủ yếu từ protein (chiếm tới 80-85%) và RNA ribosome (rRNA). Chức năng chính của nhân con là tổng hợp ribosome. Đây là nơi các gen mã hóa cho rRNA được phiên mã một cách tích cực để tạo ra các phân tử rRNA. Đồng thời, các protein ribosome, được tổng hợp trong tế bào chất và nhập vào nhân, sẽ được vận chuyển đến nhân con. Tại đây, các phân tử rRNA và protein ribosome sẽ được lắp ráp lại với nhau để tạo thành các tiểu phần lớn và tiểu phần nhỏ của ribosome. Sau khi được lắp ráp hoàn chỉnh, các tiểu phần này sẽ được xuất ra tế bào chất qua các lỗ nhân để tham gia vào quá trình tổng hợp protein. Ngoài ra, nhân con còn được cho là có vai trò trong các hoạt động khác như sửa chữa tổn thương DNA và điều chỉnh chu kỳ tế bào.
Tế bào chất (Cytoplasm)
Tế bào chất là toàn bộ phần vật chất nằm giữa màng sinh chất và nhân tế bào. Tế bào chất là một hệ thống keo phức tạp, chứa đựng các bào quan và là nơi diễn ra phần lớn các hoạt động sống của tế bào.
Bào tương (Cytosol)
Bào tương là một thành phần cực kỳ quan trọng và năng động của tế bào thực vật. Đây chính là phần chất nền dạng lỏng, giống như gel, lấp đầy không gian bên trong tế bào, bao quanh nhân và các bào quan khác. Bào tương không phải là một dung dịch lỏng đơn thuần mà là một hệ keo phức tạp. Trạng thái của nó có thể thay đổi linh hoạt giữa dạng lỏng (sol) và dạng đặc hơn (gel) tùy thuộc vào điều kiện sinh lý của tế bào, nhiệt độ, pH và nồng độ ion.
- Trạng thái Sol (Solution): Là trạng thái lỏng, chảy. Trong trạng thái này, các phân tử protein và các thành phần khác trong bào tương phân tán tương đối xa nhau và có ít liên kết hơn. Bào tương ở dạng sol có độ nhớt thấp, cho phép các phân tử và bào quan di chuyển dễ dàng.
- Trạng thái Gel (Gelatinous): Là trạng thái đặc, giống như thạch. Trong trạng thái này, các phân tử protein (đặc biệt là các sợi của bộ khung xương tế bào như actin) hình thành một mạng lưới không gian ba chiều, liên kết chằng chịt với nhau và giữ các phân tử nước bên trong. Bào tương ở dạng gel có độ nhớt cao, cứng hơn và hạn chế sự di chuyển của các thành phần bên trong.
Sự chuyển đổi giữa trạng thái sol và gel, gọi là sự chuyển đổi sol-gel (sol-gel transition), là một quá trình thuận nghịch và được điều khiển chặt chẽ bởi tế bào. Yếu tố chính chi phối quá trình này là sự trùng hợp và giải trùng hợp của các sợi actin, một thành phần quan trọng của bộ khung xương tế bào. Khi có tác động cơ học từ bên ngoài, tế bào có thể nhanh chóng chuyển vùng bào tương sang trạng thái gel để trở nên cứng hơn và chống chịu tốt hơn.
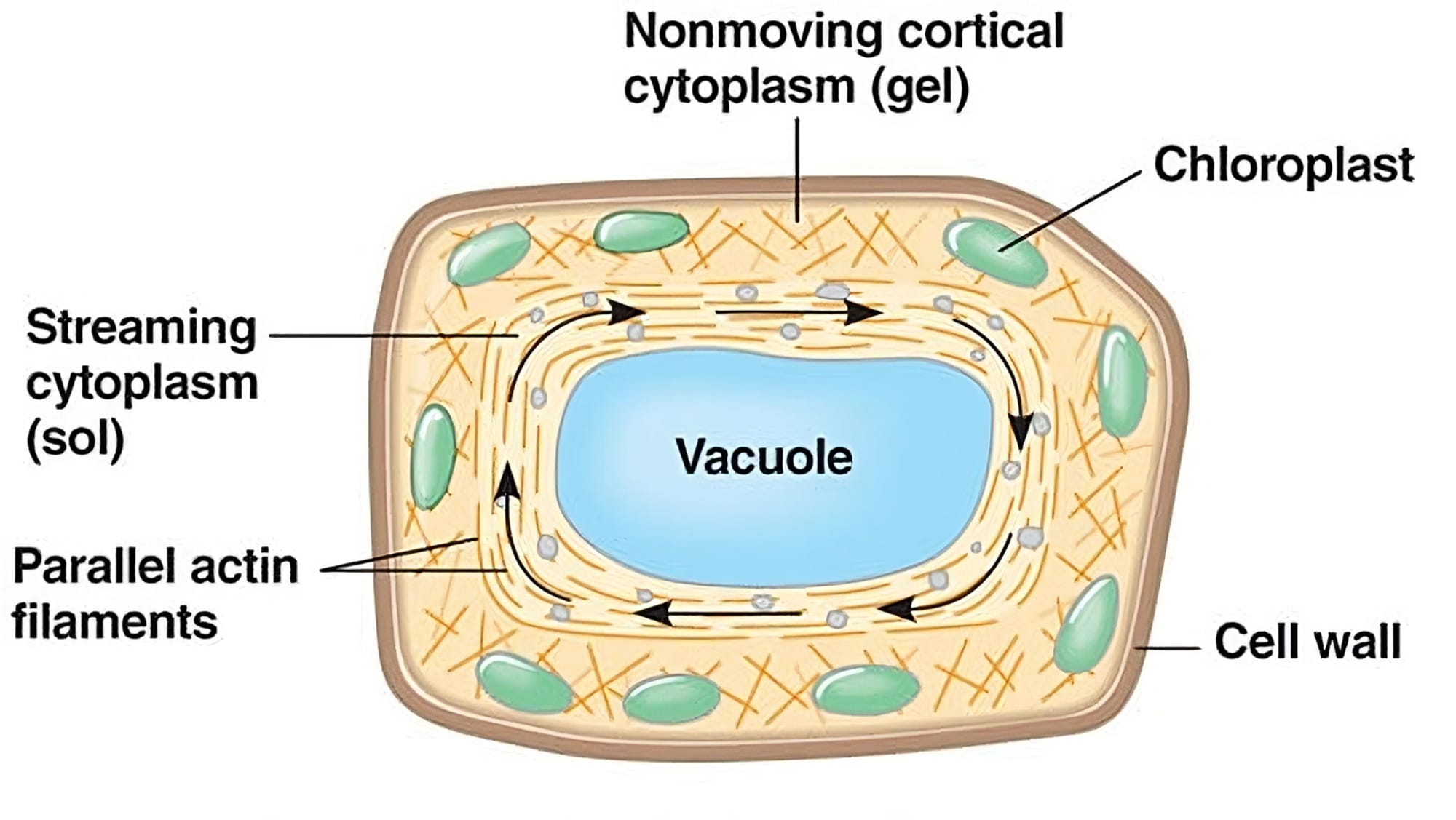
Bào tương không đứng yên mà luôn có sự chuyển động, được gọi là luồng tế bào chất (cytoplasmic streaming hay cyclosis). Sự chuyển động này giúp vận chuyển chất dinh dưỡng, các phân tử tín hiệu, bào quan và các sản phẩm trao đổi chất đến những nơi cần thiết trong tế bào, đảm bảo tế bào hoạt động hiệu quả, đặc biệt trong các tế bào thực vật lớn. Bộ khung xương tế bào nằm trong bào tương có vai trò như một "bộ xương" linh hoạt, giúp tế bào có hình dạng xác định, chống lại các tác động cơ học và là "đường ray" cho sự vận chuyển nội bào.
Bào tương không chỉ là một cái túi chứa các bào quan mà là trung tâm của nhiều hoạt động sống thiết yếu của tế bào. Rất nhiều phản ứng sinh hóa cốt lõi và quá trình truyền tín hiệu diễn ra tại đây.
Bộ khung tế bào (Cytoskeleton)
Bộ khung tế bào trong tế bào thực vật là một hệ thống cực kỳ năng động và phức tạp, đóng vai trò như "bộ xương và hệ thống cơ bắp" nội bào. Không giống như bộ xương của động vật, nó không phải là một cấu trúc tĩnh mà là một mạng lưới các sợi protein có khả năng lắp ráp, tháo dỡ và tổ chức lại một cách linh hoạt để đáp ứng các nhu cầu của tế bào.
Bộ khung tế bào thực vật được cấu tạo từ ba loại sợi chính:
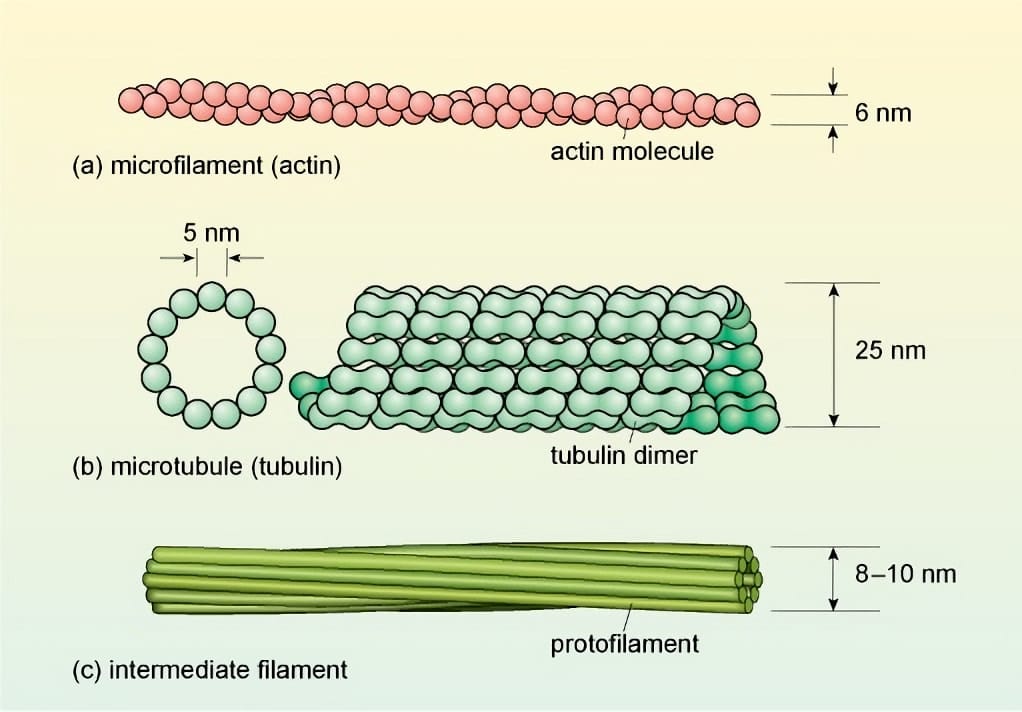
- Vi sợi (Microfilaments) là những sợi mảnh nhất trong bộ khung tế bào (đường kính khoảng 6-7 nm). Chúng được tạo thành từ sự trùng hợp của các protein hình cầu có tên là actin. Hai chuỗi actin xoắn lại với nhau tạo thành một sợi vi. Vi sợi có tính phân cực, gồm một đầu cộng (phát triển nhanh) và một đầu trừ (phát triển chậm). Các vi sợi actin tạo thành một "đường ray" vòng quanh chu vi tế bào. Các bào quan (như lục lạp, ty thể) gắn với protein vận động myosin sẽ "trượt" dọc theo đường ray actin này, tạo ra một dòng chảy tuần hoàn của bào tương. Luồng chảy này giúp phân phối đều các chất dinh dưỡng, bào quan và các phân tử tín hiệu, đặc biệt quan trọng trong các tế bào thực vật lớn. Mạng lưới vi sợi ở vùng vỏ tế bào (cortical actin array) ảnh hưởng đến sự vận chuyển và lắng đọng của các túi Golgi chứa vật liệu xây dựng thành tế bào. Vi sợi actin hình thành nên vòng thắt trong giai đoạn phân chia tế bào chất ở một số loài tảo, và tham gia vào việc định vị mặt phẳng phân chia.
- Vi ống (Microtubules) là những sợi rỗng, cứng và lớn nhất trong bộ khung tế bào (đường kính khoảng 25 nm). Chúng được tạo thành từ sự trùng hợp của các protein hình cầu gọi là tubulin (gồm α-tubulin và β-tubulin). Các tubulin kết hợp thành các chuỗi dài (protofilament), và 13 chuỗi như vậy xếp song song tạo thành một ống rỗng. Vi ống cũng có tính phân cực (đầu cộng và đầu trừ). Ở tế bào thực vật, các vi ống nằm ở vùng vỏ (ngay dưới màng sinh chất) tạo thành một cấu trúc gọi là mạng vi ống vỏ (cortical microtubule array). Các vi ống này hoạt động như một "khuôn mẫu", định hướng cho sự di chuyển của các phức hợp enzyme tổng hợp cellulose trên màng sinh chất. Do đó, hướng của các vi ống sẽ quyết định hướng sắp xếp của các sợi cellulose trong thành tế bào. Tế bào sẽ lớn lên theo hướng vuông góc với hướng của các sợi cellulose. Đây là cơ chế then chốt kiểm soát hình dạng cuối cùng của tế bào thực vật. Vi ống là thành phần chính tạo nên thoi phân bào (spindle), có nhiệm vụ phân chia chính xác các nhiễm sắc thể về hai tế bào con. Ở thực vật, chúng còn hình thành nên hai cấu trúc đặc trưng là dải tiền kì (Preprophase band) và bộ máy tạo vách (Phragmoplast). Vi ống cũng là "đường ray" cho sự di chuyển của các bào quan và các túi vận chuyển, sử dụng các protein vận động như kinesin và dynein.
- Sợi trung gian (Intermediate Filaments) có kích thước trung gian giữa vi sợi và vi ống (đường kính khoảng 8-10 nm). Chúng được tạo thành từ nhiều loại protein sợi khác nhau, rất bền và ổn định. Không giống như hai loại sợi kia, sợi trung gian không có tính phân cực và ít năng động hơn. Vai trò của sợi trung gian ở thực vật vẫn chưa được hiểu rõ hoàn toàn như ở động vật. Tuy nhiên, các nghiên cứu cho thấy chúng có chức năng chịu lực cơ học và duy trì cấu trúc. Chúng tạo thành một mạng lưới bao quanh nhân và tỏa ra khắp tế bào chất, giúp neo giữ nhân và các bào quan khác vào đúng vị trí của chúng.
Một điểm khác biệt đáng chú ý là tế bào của thực vật bậc cao (thực vật hạt trần và hạt kín) không có trung thể và trung tử. Ở tế bào động vật, trung thể là trung tâm tổ chức vi ống chính, đóng vai trò cốt lõi trong việc hình thành thoi phân bào trong quá trình nguyên phân. Mặc dù thiếu vắng cấu trúc trung tâm này, tế bào thực vật vẫn có khả năng tổ chức các vi ống để hình thành một thoi phân bào hoàn chỉnh và chức năng, đảm bảo sự phân chia chính xác của các nhiễm sắc thể về hai tế bào con. Trong khi động vật tiến hóa một giải pháp tập trung dựa trên trung thể, thực vật bậc cao lại phát triển các cơ chế khác, có thể phân tán hơn, để tập hợp các vi ống thành thoi phân bào, có thể bắt nguồn từ các vị trí trên bề mặt màng nhân hoặc các trung tâm tổ chức khác trong tế bào chất. Đây là một ví dụ kinh điển về sự tiến hóa hội tụ chức năng: cùng một mục tiêu (phân chia nhiễm sắc thể) có thể đạt được bằng các con đường phân tử và cấu trúc khác nhau. Nó thách thức quan niệm rằng một cấu trúc cụ thể là "bắt buộc" cho một chức năng cơ bản, thay vào đó cho thấy sự đa dạng và sáng tạo của các giải pháp mà sự sống đã phát triển.
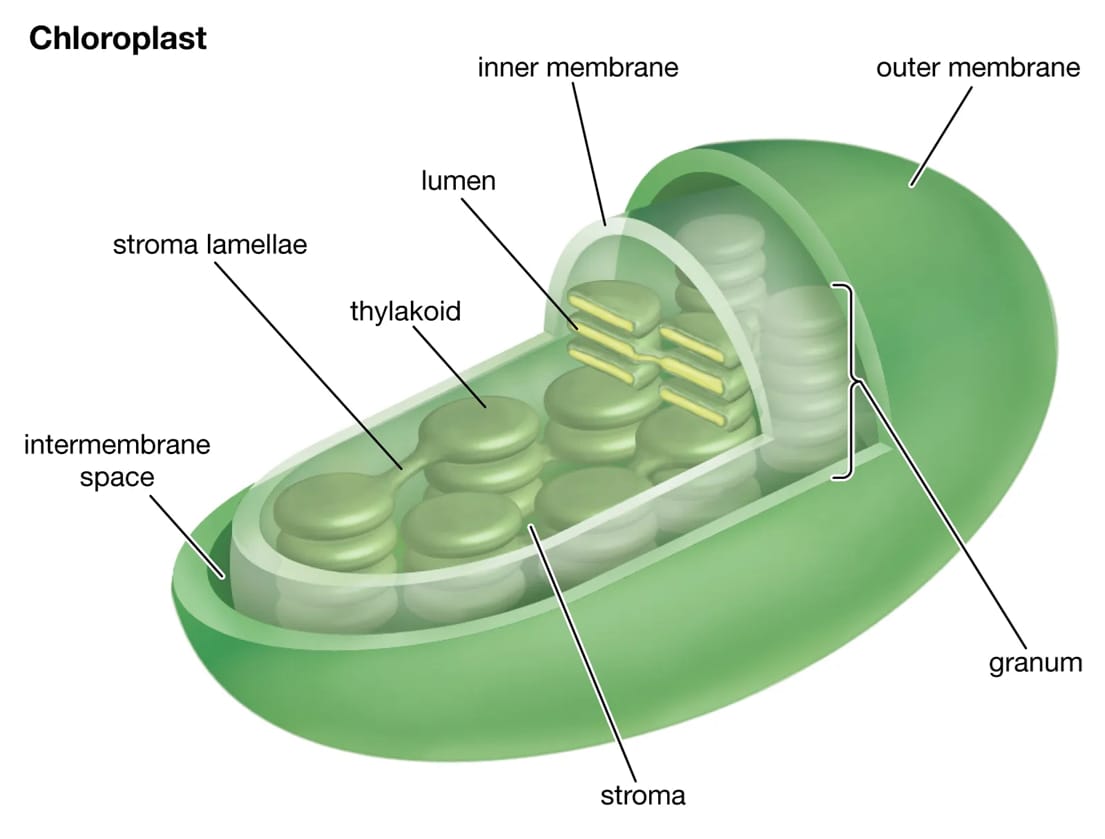
Lục lạp (Chloroplast)
Lục lạp là bào quan đặc trưng và có lẽ là quan trọng nhất của tế bào thực vật, là nền tảng cho phương thức sống tự dưỡng của toàn bộ giới thực vật. Chúng chỉ được tìm thấy ở các bộ phận có màu xanh của cây, đặc biệt nhiều ở các tế bào mô giậu của lá. Chức năng duy nhất của lục lạp là thực hiện quá trình quang hợp: sử dụng năng lượng ánh sáng, nước và khí CO2 để tổng hợp nên các hợp chất hữu cơ và giải phóng khí O2.
Xem thêm tại bài: Quang hợp.
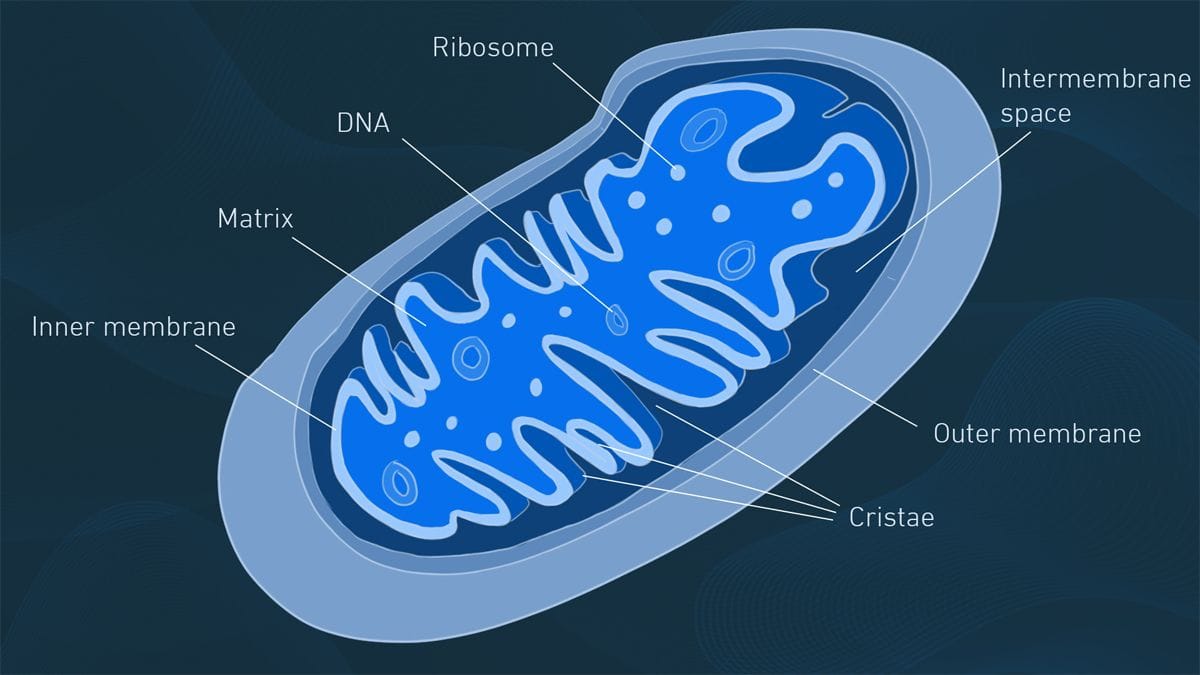
Ty thể (Mitochondria)
Nếu lục lạp là nhà máy sản xuất, thì ty thể chính là nhà máy điện của tế bào. Ty thể có mặt trong hầu hết các tế bào nhân thực, bao gồm cả tế bào thực vật và động vật. Chúng thực hiện quá trình hô hấp tế bào, một chuỗi các phản ứng oxy hóa các phân tử hữu cơ (như glucose) để giải phóng năng lượng được lưu trữ trong đó và tổng hợp nên adenosine triphosphate (ATP) - "đồng tiền năng lượng" chung cho mọi hoạt động sống của tế bào. Ở thực vật, ty thể đặc biệt quan trọng vào ban đêm hoặc ở các mô không quang hợp (như rễ), khi năng lượng từ quang hợp không có sẵn.
Xem thêm tại bài: Hô hấp tế bào.
Lưới nội chất (Endoplasmic Reticulum - ER)
Lưới nội chất là một mạng lưới rộng lớn gồm các túi màng dẹt (gọi là bể chứa) và các ống màng thông với nhau, lan tỏa khắp tế bào chất. Khoang bên trong của lưới nội chất, gọi là xoang ER, là một không gian riêng biệt. Màng ER liên tục với màng ngoài của nhân, tạo thành một hệ thống không gian thông suốt. Có hai vùng của lưới nội chất có cấu trúc và chức năng khác nhau:
Lưới nội chất hạt (Rough ER): Bề mặt phía tế bào chất của màng có đính rất nhiều ribosome, tạo ra vẻ ngoài "sần sùi" hay "có hạt" dưới kính hiển vi điện tử. Chức năng chính của lưới nội chất hạt là tổng hợp protein. Các chuỗi polypeptide được tổng hợp bởi các ribosome này sẽ được đưa trực tiếp vào xoang ER. Tại đây, chúng được cuộn gập thành cấu trúc không gian ba chiều chính xác và trải qua các biến đổi ban đầu, chẳng hạn như quá trình glycosyl hóa (gắn thêm các chuỗi carbohydrate ngắn). Các protein được tổng hợp ở đây chủ yếu là các protein sẽ được tiết ra khỏi tế bào, các protein của màng sinh chất và các protein của các bào quan khác trong hệ thống nội màng.
Lưới nội chất trơn (Smooth ER): Bề mặt màng không có ribosome đính vào. Lưới nội chất trơn có các chức năng đa dạng và chuyên biệt hơn: tổng hợp lipid, chuyển hóa carbonhydrate, giải độc tế bào, dự trữ ion canxi...
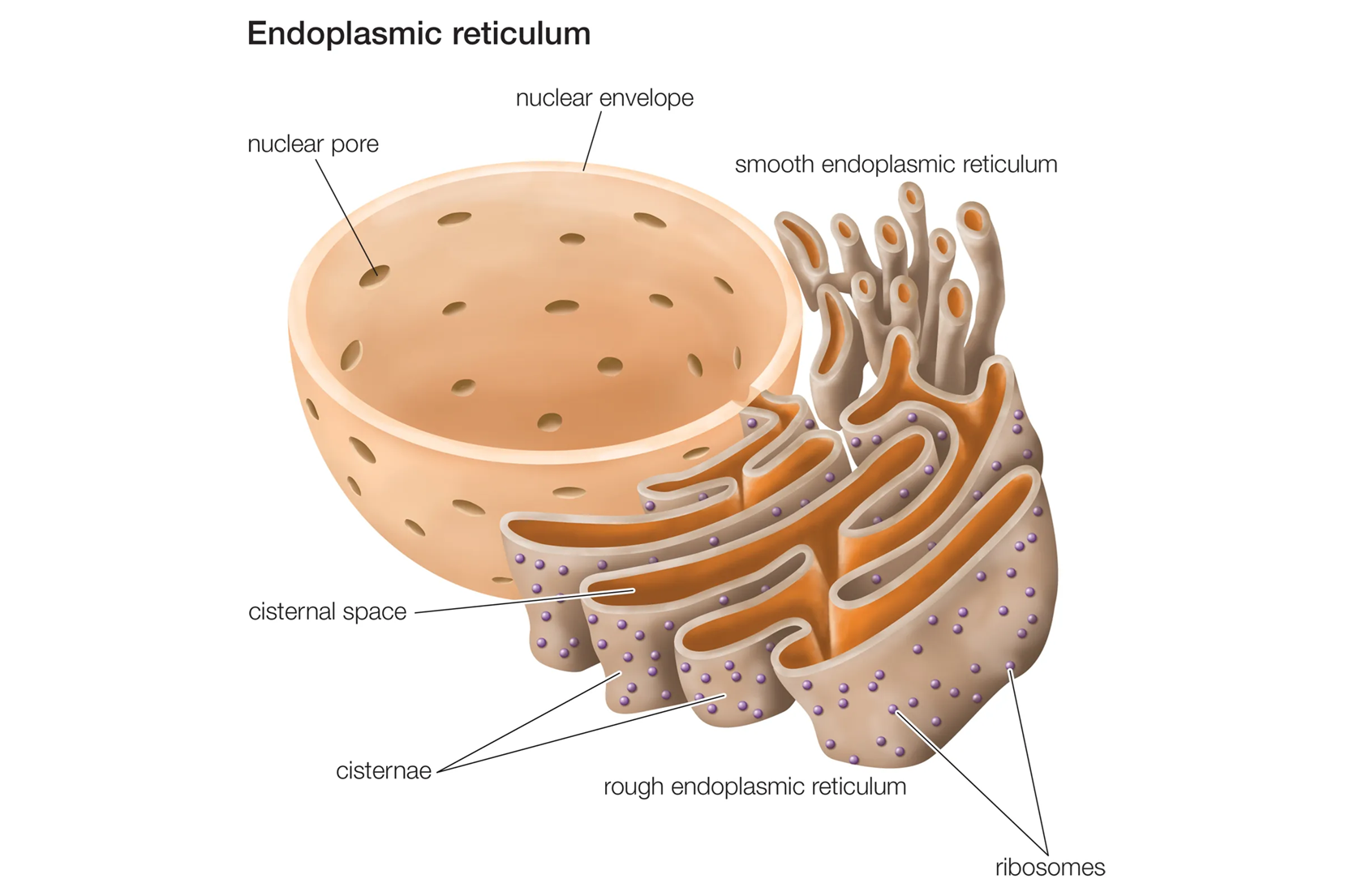
Mạng lưới ER của các tế bào thực vật không bị cô lập mà kết nối với nhau thông qua các kênh xuyên màng gọi là cầu sinh chất (Plasmodesmata). Một ống ER trơn nén chặt, được gọi là desmotubule, chạy xuyên qua tâm của mỗi cầu sinh chất, tạo ra một sự liên tục vật lý của hệ thống ER giữa các tế bào liền kề. Điều này cho phép sự vận chuyển trực tiếp các phân tử tín hiệu, protein, và thậm chí cả virus từ tế bào này sang tế bào khác, hợp nhất các tế bào riêng lẻ thành một thể thống nhất gọi là hợp bào (symplast).
Bộ máy Golgi (Golgi Apparatus)
Sau khi được tổng hợp và xử lý sơ bộ tại lưới nội chất, nhiều protein và lipid sẽ được vận chuyển đến bộ máy Golgi để tiếp tục được hoàn thiện và phân phối. Bộ máy Golgi bao gồm một chồng các túi màng dẹt, không thông nhau, được gọi là các bể (cisternae).
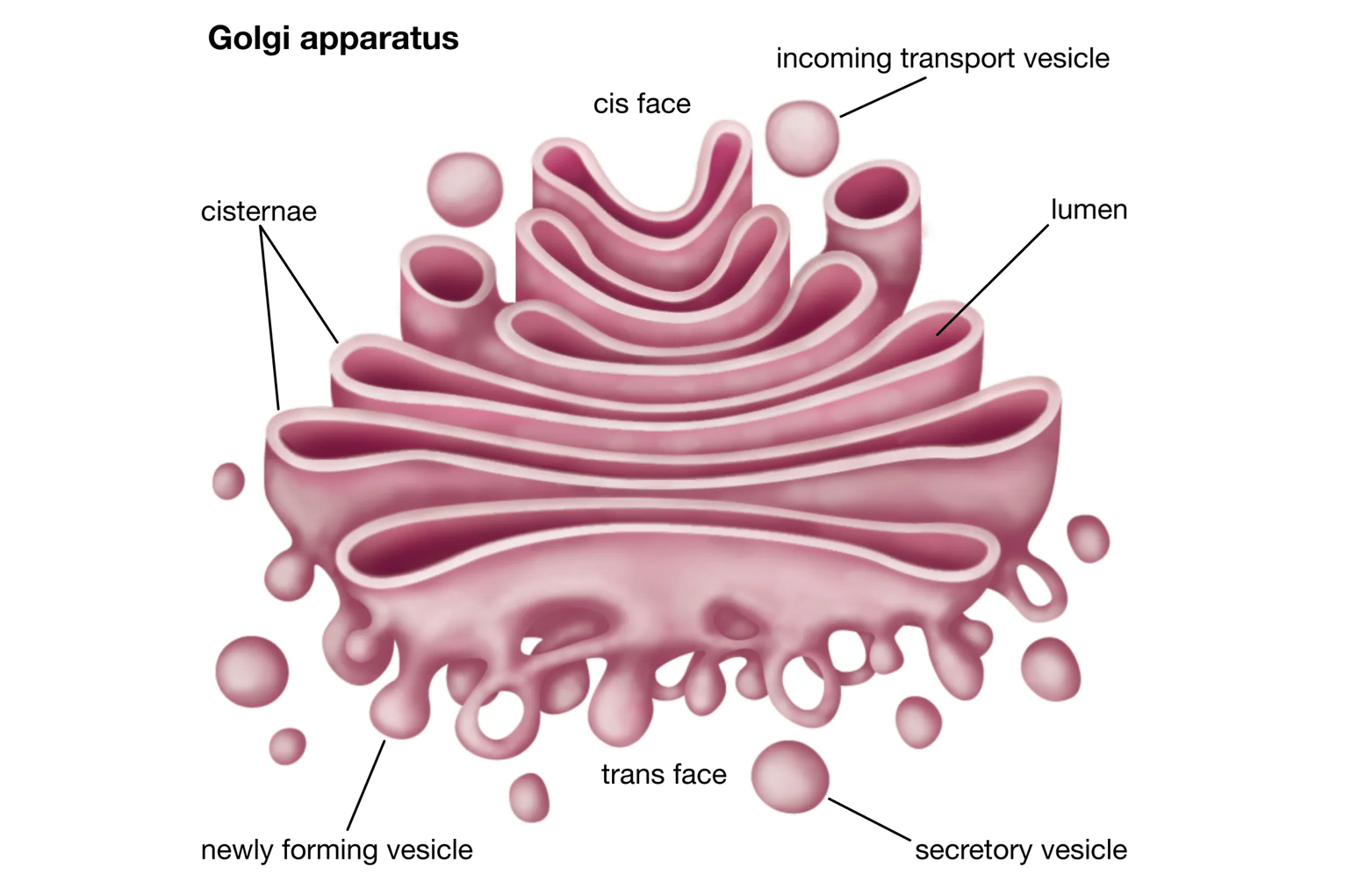
Bộ máy Golgi hoạt động như một "trung tâm xử lý và phân phối" hay "bưu điện" của tế bào. Nó có một mặt nhận (mặt cis) hướng về phía lưới nội chất và một mặt xuất (mặt trans) hướng về phía màng sinh chất. Các túi vận chuyển nhỏ tách ra từ ER, mang theo protein và lipid, sẽ dung hợp với mặt cis của Golgi. Khi các phân tử này di chuyển qua các bể của Golgi từ mặt cis đến mặt trans, chúng sẽ được tiếp tục biến đổi, ví dụ như hoàn thiện quá trình glycosyl hóa, cắt bớt một số đoạn polypeptide, hoặc gắn thêm các nhóm chức khác. Cuối cùng, tại mặt trans, các sản phẩm đã hoàn thiện sẽ được phân loại, "dán nhãn" hóa học và đóng gói vào các túi vận chuyển mới. Các túi này sau đó sẽ tách ra và di chuyển đến các đích cuối cùng của chúng, có thể là màng sinh chất (để xuất bào), không bào, hoặc các vị trí khác trong tế bào.
Ribosome
Ribosome là những cỗ máy phân tử cực nhỏ nhưng có số lượng rất lớn trong tế bào, chịu trách nhiệm cho một trong những quá trình cơ bản nhất của sự sống: tổng hợp protein. Chúng không có màng bao bọc và được cấu tạo từ RNA ribosome (rRNA) và protein. Mỗi ribosome bao gồm hai tiểu phần, một tiểu phần lớn và một tiểu phần nhỏ, chỉ kết hợp với nhau khi tiến hành dịch mã.
Chức năng của ribosome là "đọc" thông tin di truyền được mã hóa trong phân tử RNA thông tin (mRNA) và sử dụng thông tin đó để lắp ráp các axít amin thành một chuỗi polypeptide theo đúng trình tự. Quá trình này được gọi là dịch mã. Ribosome có thể được tìm thấy ở hai vị trí trong tế bào chất, tương ứng với hai con đường tổng hợp protein khác nhau:
- Ribosome tự do: Trôi nổi trong dịch bào, chúng tổng hợp các protein sẽ hoạt động ngay trong tế bào chất, trong nhân, ty thể hoặc lục lạp.
- Ribosome liên kết: Tạm thời bám vào màng của lưới nội chất hạt, chúng tổng hợp các protein được định sẵn để xuất ra khỏi tế bào hoặc để trở thành một phần của hệ thống nội màng.
Không bào trung tâm (Central Vacuole)
Một trong những đặc điểm dễ nhận biết nhất của một tế bào thực vật trưởng thành là sự hiện diện của một không bào trung tâm rất lớn. Bào quan này có thể chiếm 30-90% thể tích của tế bào, đẩy tế bào chất và nhân ép sát vào vách tế bào.
Không bào là một túi lớn được bao bọc bởi một lớp màng duy nhất gọi là màng không bào (tonoplast). Màng tonoplast có tính thấm chọn lọc, kiểm soát chặt chẽ sự di chuyển của các chất giữa tế bào chất và khoang bên trong không bào. Khoang này chứa một dung dịch nước gọi là dịch tế bào (cell sap). Thành phần của dịch tế bào rất đa dạng, bao gồm nước, các ion vô cơ (như K+, Na+, Cl−, Ca2+), đường, axít hữu cơ, axít amin, protein, các sắc tố tan trong nước (tạo ra màu đỏ, tím, xanh cho nhiều loại hoa, quả và lá), các chất chuyển hóa thứ cấp (như nicotine, caffeine, tannin), enzyme thủy phân, và cả các sản phẩm phụ hoặc chất thải của quá trình trao đổi chất.
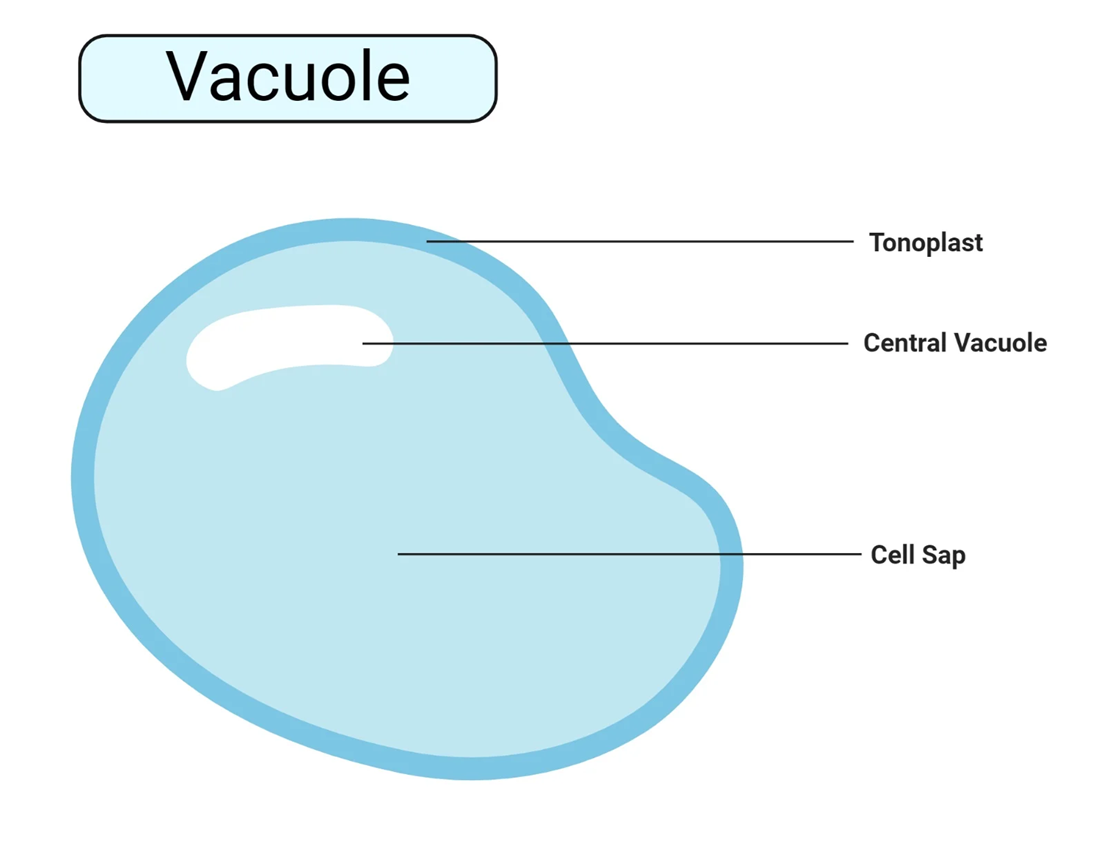
Không bào trung tâm không phải là một không gian chết mà là một bào quan đa chức năng, đóng vai trò sống còn đối với tế bào thực vật: dự trữ dinh dưỡng, duy trì áp suất trương nước, tăng trưởng tế bào, phân giải bào quan già cỗi hoặc các đại phân tử.
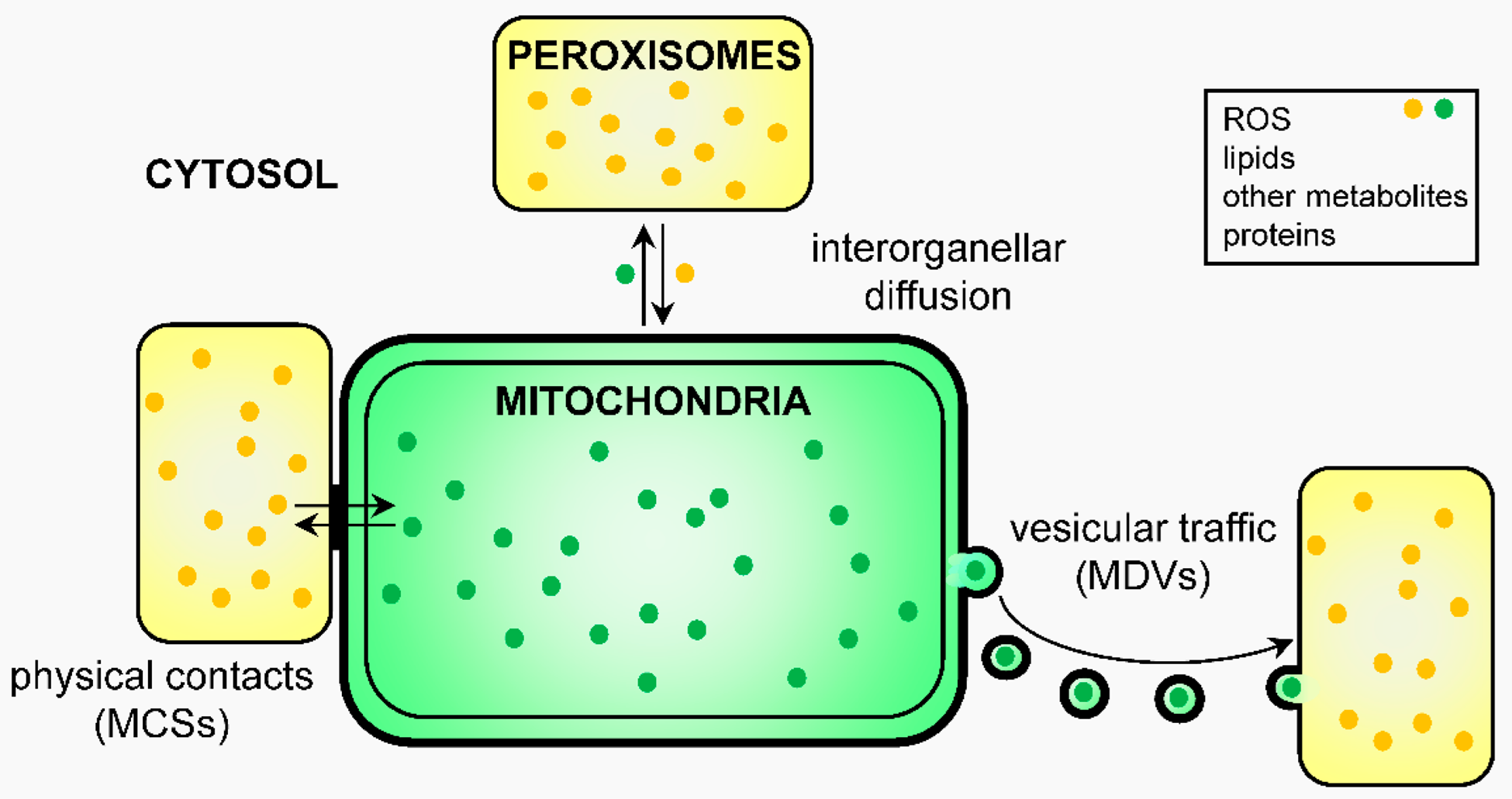
Peroxisome
Peroxisome là những bào quan nhỏ, hình cầu, được bao bọc bởi một lớp màng đơn. Chúng chứa các enzyme chuyên biệt tham gia vào nhiều quá trình trao đổi chất khác nhau. Một chức năng đặc trưng của peroxisome là sử dụng oxy để phân giải các phân tử phức tạp. Trong quá trình này, chúng tạo ra một sản phẩm phụ độc hại là hydrogen peroxide (H2O2). Tuy nhiên, peroxisome cũng chứa một lượng lớn enzyme catalase, có khả năng phân giải nhanh chóng H2O2 thành nước và oxy, hai sản phẩm vô hại. Các chức năng cụ thể của peroxisome ở thực vật bao gồm phân giải axít béo thành các phân tử nhỏ hơn để cung cấp năng lượng cho hạt nảy mầm và tham gia vào quá trình quang hô hấp ở lá.
Tế bào như một hệ thống tích hợp
Sau khi đã phân tích chi tiết từng thành phần riêng lẻ, điều quan trọng là phải tổng hợp các kiến thức này để xây dựng một bức tranh toàn cảnh về tế bào thực vật như một hệ thống sống, năng động và có sự phối hợp chặt chẽ. Tế bào không phải là một tập hợp ngẫu nhiên các bào quan, mà là một cỗ máy sinh học phức tạp, trong đó mỗi bộ phận tương tác và giao tiếp với nhau để thực hiện các chức năng ở cấp độ cao hơn.
Để hiểu rõ sự phối hợp giữa các bào quan, chúng ta có thể xem xét một quy trình cụ thể: con đường tổng hợp và vận chuyển một protein được tiết ra ngoài tế bào (ví dụ như một enzyme tiêu hóa được rễ tiết ra). Quá trình này là một chuỗi các sự kiện liên tiếp, thể hiện sự hợp tác hoàn hảo của hệ thống nội màng và nhân tế bào.
Mọi thứ bắt đầu từ nhân. Tại đây, thông tin di truyền mã hóa cho protein cần tổng hợp được sao chép từ một đoạn DNA (gen) thành một phân tử RNA thông tin (mRNA) → Phân tử mRNA sau khi được xử lý hoàn chỉnh sẽ rời khỏi nhân qua các phức hợp lỗ nhân và đi ra tế bào chất → Trong tế bào chất, một ribosome sẽ bám vào phân tử mRNA và bắt đầu quá trình dịch mã. Vì đây là một protein tiết, chuỗi polypeptide mới sinh sẽ có một "tín hiệu" đặc biệt ở đầu, giúp toàn bộ phức hợp ribosome-mRNA được hướng đến và bám vào màng của lưới nội chất hạt → Chuỗi polypeptide sau đó được tổng hợp và đưa trực tiếp vào xoang của lưới nội chất → Bên trong xoang ER, chuỗi polypeptide sẽ cuộn gập thành cấu trúc không gian ba chiều chính xác và có thể trải qua các biến đổi hóa học ban đầu, như được gắn thêm các chuỗi carbohydrate ngắn → Các protein đã được biến đổi sẽ được bao bọc trong các túi vận chuyển nhỏ, tách ra từ màng ER và di chuyển đến bộ máy Golgi → Các túi vận chuyển dung hợp với mặt cis của Golgi. Khi protein di chuyển qua các bể của Golgi, nó sẽ được tiếp tục biến đổi và hoàn thiện → Tại mặt trans, protein đã hoàn chỉnh sẽ được phân loại và đóng gói vào các túi tiết → Các túi tiết di chuyển dọc theo các sợi của bộ khung tế bào đến màng sinh chất → Tại đây, màng của túi tiết sẽ dung hợp với màng sinh chất, giải phóng protein ra môi trường bên ngoài tế bào. Quá trình này được gọi là xuất bào.
Con đường này cho thấy rõ rằng không một bào quan nào có thể tự mình hoàn thành nhiệm vụ. Chức năng "tổng hợp và tiết protein" không thuộc về nhân, ribosome, ER hay Golgi một cách riêng lẻ. Thay vào đó, chức năng này nó chỉ "nổi lên" khi tất cả các bào quan này hoạt động phối hợp với nhau theo một trình tự không gian và thời gian được điều khiển chặt chẽ. Điều này minh họa một nguyên tắc cơ bản của sinh học hệ thống: để hiểu được các chức năng phức tạp của tế bào, việc nghiên cứu từng bộ phận là cần thiết nhưng chưa đủ; cần phải xem xét các mạng lưới tương tác và các dòng chảy thông tin, vật chất, năng lượng giữa các thành phần đó.
Sự phối hợp không chỉ diễn ra bên trong một tế bào mà còn lan rộng ra toàn bộ cơ thể thực vật. Khác với tế bào động vật thường được ngăn cách bởi không gian gian bào, các tế bào thực vật được kết nối trực tiếp với nhau thông qua hàng ngàn kênh giao tiếp nhỏ gọi là cầu sinh chất. Cầu sinh chất là những ống hẹp, được lót bởi màng sinh chất, xuyên qua vách tế bào để nối liền tế bào chất của các tế bào lân cận. Cấu trúc này tạo ra một thể liên tục của tế bào chất trên toàn bộ cơ thể thực vật, được gọi là thể liên bào (symplast). Thông qua mạng lưới symplast này, các phân tử nhỏ như nước, ion, đường, axít amin, và thậm chí cả các phân tử tín hiệu lớn hơn như protein và RNA, có thể di chuyển trực tiếp từ tế bào này sang tế bào khác mà không cần phải vượt qua rào cản của màng sinh chất và vách tế bào.
Cầu sinh chất đóng vai trò thiết yếu trong việc vận chuyển chất dinh dưỡng (ví dụ, đường được sản xuất ở lá được vận chuyển đến rễ), truyền các tín hiệu phát triển (như hormone), và phối hợp các đáp ứng của cây đối với môi trường. Chúng biến một tập hợp các tế bào riêng lẻ thành một cơ thể thực vật thống nhất, hoạt động một cách có tổ chức và đồng bộ.
Sự tồn tại của thể liên bào thông qua cầu sinh chất mang một ý nghĩa sâu sắc, định nghĩa lại khái niệm cá thể ở thực vật. Nếu ở động vật, tế bào thường được xem là đơn vị cơ bản, độc lập, giao tiếp với nhau chủ yếu qua các tín hiệu ngoại bào, thì ở thực vật, ranh giới giữa các tế bào trở nên mờ nhạt hơn. Môi trường nội bào của một tế bào ở lá có thể liên thông trực tiếp với một tế bào ở rễ. Các tín hiệu phát triển, các yếu tố điều hòa, và thậm chí cả virus thực vật, có thể lan truyền khắp cơ thể thông qua mạng lưới liên bào này. Do đó, có thể xem toàn bộ cơ thể thực vật không chỉ là một tập hợp hàng tỷ tế bào, mà còn là một "siêu tế bào" (supracellular organism), trong đó các thành tế bào chỉ đóng vai trò như những vách ngăn phân chia một không gian tế bào chất chung. Đây là một sự khác biệt triệt để và cơ bản trong triết lý tổ chức cơ thể so với giới động vật.
Kết luận
Qua phân tích chi tiết, có thể thấy rằng tế bào thực vật điển hình không phải là một cấu trúc đơn giản mà là một hệ thống cực kỳ phức tạp và được tổ chức một cách tinh vi. Từ pháo đài bảo vệ là thành tế bào, cánh cổng kiểm soát năng động là màng sinh chất, đến trung tâm chỉ huy là nhân, và các nhà máy năng lượng, sản xuất chuyên biệt bên trong tế bào chất, mỗi thành phần đều có một cấu trúc siêu vi và thành phần hóa học được tối ưu hóa cho chức năng của nó.
Quan trọng hơn, các cấu trúc này không tồn tại một cách cô lập. Chúng là một phần của một hệ thống tích hợp cao độ, trong đó hoạt động của mỗi bào quan được phối hợp nhịp nhàng với các bào quan khác thông qua các dòng chảy vật chất, năng lượng và thông tin. Con đường tổng hợp protein tiết là một minh chứng rõ ràng cho sự phối hợp này, cho thấy cách thông tin từ nhân được chuyển hóa thành sản phẩm vật chất thông qua một dây chuyền sản xuất liên bào quan. Hơn nữa, sự tích hợp này không chỉ giới hạn trong một tế bào mà còn mở rộng ra toàn bộ cơ thể thông qua mạng lưới cầu sinh chất, biến cơ thể thực vật thành một thể thống nhất về mặt chức năng.
Cấu trúc độc đáo của tế bào thực vật với thành cellulose, lục lạp và không bào trung tâm là kết quả của hàng triệu năm tiến hóa, một sự thích nghi hoàn hảo cho lối sống tự dưỡng và cố định. Nó đã tạo ra một cỗ máy sinh học hiệu quả, có khả năng khai thác nguồn năng lượng vô tận từ mặt trời, xây dựng nên những cấu trúc phức tạp từ các chất vô cơ đơn giản, và qua đó, tạo thành nền tảng cho hầu hết các hệ sinh thái trên Trái Đất. Do đó, việc nghiên cứu tế bào thực vật không chỉ là khám phá đơn vị cơ bản của sự sống thực vật mà còn là chiêm ngưỡng một trong những minh chứng hùng hồn nhất cho sự đa dạng, phức tạp và tinh vi của các giải pháp mà sự sống đã phát triển để tồn tại và phát triển mạnh mẽ.


