
Hô hấp
Đường phân, Krebs, chuỗi truyền điện tử
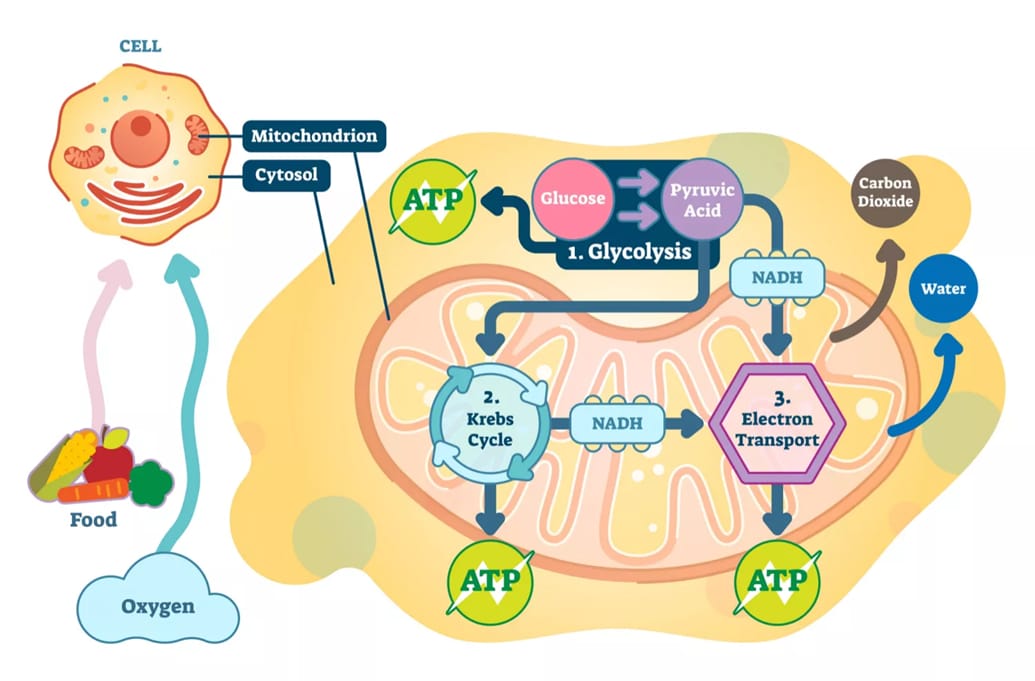
Hô hấp tế bào là một quá trình trao đổi chất nền tảng, giữ vai trò trung tâm trong đời sống của mọi sinh vật nhân thực, bao gồm cả thực vật. Đây là chuỗi các phản ứng sinh hóa phức tạp nhằm chuyển đổi năng lượng hóa học được lưu trữ trong các phân tử hữu cơ - chủ yếu là glucose được tạo ra từ quá trình quang hợp - thành adenosine triphosphate (ATP), dạng năng lượng phổ quát mà tế bào có thể sử dụng trực tiếp cho mọi hoạt động sống. Bào quan chịu trách nhiệm chính cho quá trình này là ty thể, thường được mệnh danh là "nhà máy năng lượng của tế bào".
Cấu tạo ty thể
Ty thể là các bào quan có mặt trong tế bào chất của tất cả các sinh vật nhân thực, với hình dạng và kích thước đa dạng, có thể là hình cầu, hình que hoặc dạng sợi dài. Kích thước của chúng thường dao động với đường kính từ 0,5-1 µm và chiều dài có thể lên tới 7 µm. Những cấu trúc này không tĩnh tại mà rất năng động, liên tục di chuyển, hợp nhất và phân chia bên trong tế bào.
Cấu trúc của ty thể được đặc trưng bởi một hệ thống màng kép, tạo ra các khoang riêng biệt với những chức năng chuyên hóa cao. Màng ngoài ty thể là một lớp màng trơn, không gấp khúc, có tính thấm cao. Đặc tính này có được là nhờ sự hiện diện của các protein kênh gọi là porin, cho phép các phân tử nhỏ (như pyruvate) và một số protein di chuyển tự do từ tế bào chất vào không gian giữa hai màng. Màng trong ty thể có tính thấm chọn lọc rất cao và được gấp nếp mạnh mẽ. Đây là nơi diễn ra các quá trình quan trọng nhất của hô hấp tế bào, bao gồm chuỗi chuyền electron và tổng hợp ATP. Màng trong có hàm lượng protein rất cao, chiếm tới 65-70% khối lượng khô, chủ yếu là các enzyme và phức hợp protein của chuỗi chuyền điện tử (electron). Các lớp màng này phân định các khoang chức năng bên trong ty thể:
- Xoang gian màng là khoảng không gian hẹp giữa màng ngoài và màng trong. Đây là nơi tích tụ các ion hydro (H+) được bơm ra từ chất nền trong quá trình chuyền electron, tạo ra một gradient điện hóa cần thiết cho việc tổng hợp ATP.
- Chất nền ty thể là khoang trong cùng của ty thể, chứa một hỗn hợp đậm đặc gồm enzyme, DNA ty thể (mtDNA), ribosome, và các chất chuyển hóa khác. Đây là nơi diễn ra chu trình Krebs và quá trình oxy hóa pyruvate.
- Mào (cristae) là các nếp gấp đặc trưng của màng trong, có vai trò làm tăng đáng kể diện tích bề mặt của màng. Cấu trúc này cho thấy một mức độ chuyên môn hóa cao độ. Sự gấp nếp không phải là ngẫu nhiên; nó phục vụ mục đích thiết yếu là tối đa hóa không gian để gắn các phức hợp protein của chuỗi chuyền electron và enzyme ATP synthase. Việc gia tăng diện tích cho phép chứa một số lượng lớn hơn các đơn vị chức năng này, trực tiếp khuếch đại khả năng sản xuất ATP của ty thể, biến cấu trúc của nó thành một bộ máy sinh hóa được tối ưu hóa cho hiệu suất năng lượng tối đa.
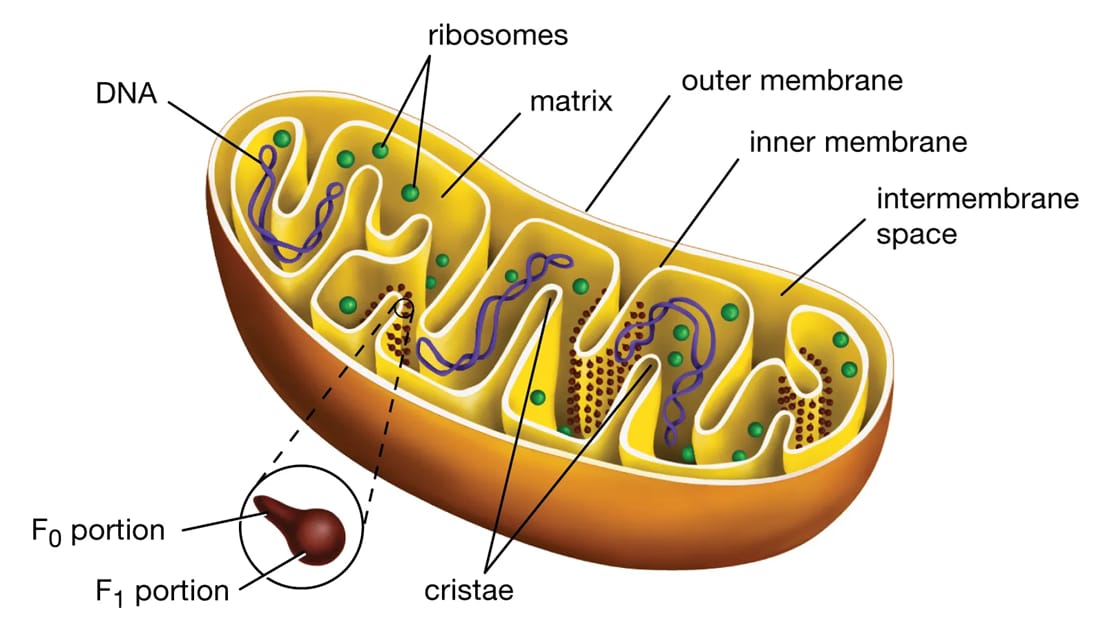
Một đặc điểm nổi bật của ty thể là chúng sở hữu hệ gen riêng, độc lập với DNA trong nhân tế bào. DNA ty thể (mtDNA) ở thực vật thường có dạng vòng, mã hóa cho một phần nhỏ các protein cần thiết cho chức năng của ty thể, chủ yếu là các tiểu đơn vị của chuỗi chuyền electron và ATP synthase, cùng với các RNA ribosome và RNA vận chuyển riêng. Sự tồn tại của hệ gen riêng này là một trong những bằng chứng mạnh mẽ nhất cho thuyết nội cộng sinh, giả thuyết cho rằng ty thể có nguồn gốc từ một vi khuẩn hiếu khí cổ đại bị một tế bào chủ sơ khai nuốt phải. Thay vì bị tiêu hóa, vi khuẩn này đã thiết lập một mối quan hệ cộng sinh, dần dần tiến hóa thành một bào quan không thể thiếu. Do có hệ gen và bộ máy tổng hợp protein riêng (ribosome, RNA), ty thể được coi là bán tự trị. Chúng có thể tự nhân đôi bằng cách phân chia, độc lập với chu kỳ của tế bào. Tuy nhiên, phần lớn các protein của ty thể (hơn 95%) được mã hóa bởi gen trong nhân, tổng hợp trong tế bào chất và sau đó được nhập vào ty thể.
Ty thể có vòng đời ngắn, chỉ kéo dài vài ngày, và trong tế bào luôn diễn ra quá trình hình thành và phân hủy ty thể. Sự phân bố của chúng trong tế bào thường liên quan đến hệ thống khung xương tế bào, đặc biệt là các vi ống, đảm bảo chúng được đưa đến những nơi có nhu cầu năng lượng cao.
Hô hấp hiếu khí
Hô hấp hiếu khí là quá trình oxy hóa hoàn toàn các phân tử hữu cơ, phổ biến nhất là glucose, thành các sản phẩm vô cơ là carbon dioxide (CO2) và nước (H2O), đồng thời giải phóng một lượng lớn năng lượng được tích lũy dưới dạng ATP và nhiệt. Về bản chất, đây là một chuỗi các phản ứng oxy hóa-khử. Phương trình tổng quát của quá trình này là:
C6H12O6 + 6O2 → 6CO2 + 6H2O + Năng lượng (ATP + Nhiệt)
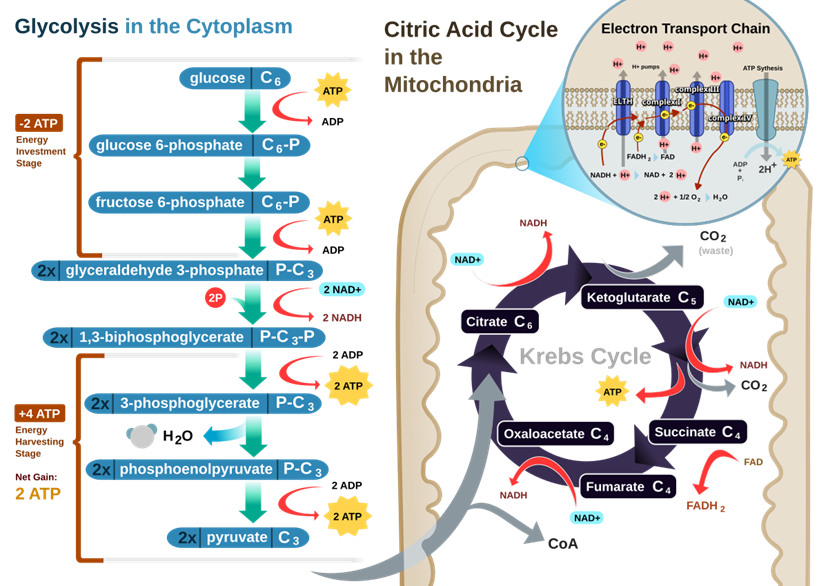
Quá trình này được chia thành ba giai đoạn chính, diễn ra ở các vị trí khác nhau trong tế bào. Đường phân là giai đoạn phân giải glucose ban đầu trong tế bào chất. Chu trình Krebs (Chu trình axít citric) oxy hóa hoàn toàn Acetyl-CoA trong chất nền ty thể. Chuỗi truyền điện tử và phosphoryl hóa xxy hóa tổng hợp ATP hiệu suất cao ở màng trong ty thể.
Đường phân
Đường phân, hay Glycolysis, là con đường trao đổi chất phổ biến nhất trong mọi sinh vật sống, đóng vai trò là điểm khởi đầu cho cả quá trình hô hấp hiếu khí và kỵ khí. Quá trình này diễn ra trong tế bào chất, không cần đến oxy và thực hiện việc phân giải một phân tử glucose (6 carbon) thành hai phân tử pyruvate (3 carbon). Song song đó, nó tạo ra các phân tử năng lượng cao là ATP và NADH.
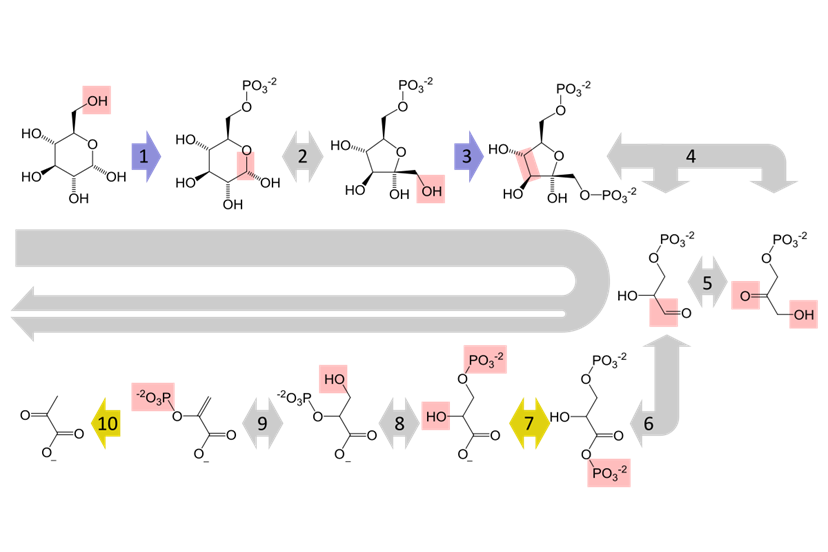
Toàn bộ con đường đường phân bao gồm 10 phản ứng enzyme, được chia thành hai giai đoạn chính:
- Giai đoạn đầu tư năng lượng: tiêu thụ ATP để biến đổi phân tử glucose thành một dạng kém ổn định hơn, sẵn sàng cho việc phân cắt.
- (1) Phân tử glucose được phosphoryl hóa (gắn thêm một nhóm phosphate) bởi enzyme Hexokinase, sử dụng một phân tử ATP, tạo thành Glucose-6-phosphate. Phản ứng này giúp giữ glucose lại bên trong tế bào.
- (2) Glucose-6-phosphate được chuyển đổi thành đồng phân của nó là Fructose-6-phosphate, nhờ enzyme Phosphoglucose Isomerase.
- (3) Một nhóm phosphate thứ hai được chuyển từ một phân tử ATP khác đến Fructose-6-phosphate, tạo thành Fructose-1,6-bisphosphate. Phản ứng này được xúc tác bởi enzyme Phosphofructokinase, một enzyme điều hòa quan trọng của quá trình đường phân.
- (4) Phân tử Fructose-1,6-bisphosphate (6 carbon) bị enzyme Aldolase cắt đôi thành hai phân tử đường 3 carbon khác nhau: Glyceraldehyde-3-phosphate (G3P) và Dihydroxyacetone phosphate (DHAP).
- (5) DHAP nhanh chóng được chuyển đổi thành G3P bởi enzyme Triose Phosphate Isomerase. Do đó, từ đây trở đi, mọi phản ứng sẽ diễn ra với hai phân tử G3P.
- Giai đoạn thu hồi năng lượng: năng lượng được giải phóng từ việc phân giải các phân tử 3 carbon và được tích trữ dưới dạng ATP và NADH. Vì giai đoạn này xử lý hai phân tử G3P (từ một phân tử glucose ban đầu), nên sản phẩm tạo ra sẽ được nhân đôi.
- (6) Mỗi phân tử Glyceraldehyde-3-phosphate bị oxy hóa. Phản ứng này giải phóng năng lượng, được thu giữ bởi NAD+ để tạo thành NADH. Phân tử đường sau đó được gắn thêm một nhóm phosphate vô cơ (Pi) để tạo thành 1,3-Bisphosphoglycerate. Phản ứng được xúc tác bởi Glyceraldehyde-3-phosphate Dehydrogenase.
- (7) Một trong các nhóm phosphate năng lượng cao của 1,3-Bisphosphoglycerate được chuyển đến ADP để tạo thành ATP. Quá trình này được gọi là phosphoryl hóa mức cơ chất, do enzyme Phosphoglycerate Kinase xúc tác, để lại phân tử 3-Phosphoglycerate.
- (8) Phân tử 3-Phosphoglycerate được sắp xếp lại, di chuyển nhóm phosphate từ vị trí carbon thứ 3 sang vị trí thứ 2, tạo thành 2-Phosphoglycerate. Enzyme xúc tác là Phosphoglycerate Mutase.
- (9) Enzyme Enolase loại bỏ một phân tử nước khỏi 2-Phosphoglycerate, tạo ra một liên kết đôi giàu năng lượng, hình thành nên phân tử Phosphoenolpyruvate (PEP).
- (10) Nhóm phosphate năng lượng cao cuối cùng được chuyển từ PEP sang ADP, tạo ra ATP. Đây cũng là một bước phosphoryl hóa mức cơ chất. Phân tử còn lại là Pyruvate, sản phẩm cuối cùng của đường phân. Phản ứng được xúc tác bởi Pyruvate Kinase.
Axít béo là một nguồn dự trữ năng lượng cực kỳ hiệu quả. Chúng không đi qua đường phân mà được phân giải thông qua một quá trình gọi là β-oxy hóa (Beta-oxidation), diễn ra bên trong chất nền của ty thể. Khi protein được sử dụng làm năng lượng (thường xảy ra khi cơ thể thiếu hụt glucose và lipid), các axít amin sẽ được phân giải. Con đường này phức tạp hơn vì có 20 loại axít amin khác nhau.
Chu trình Krebs
Chu trình Krebs, còn được gọi là chu trình axit citric hoặc chu trình axit tricarboxylic (TCA), là giai đoạn trung tâm của quá trình hô hấp tế bào. Nó diễn ra trong chất nền của ty thể ở tế bào nhân thực và là con đường chung cuối cùng cho quá trình oxy hóa carbohydrate, lipid và protein. Vai trò cốt lõi của nó là thu hoạch các electron năng lượng cao cho các phân tử mang electron là NAD+ và FAD, để tạo ra NADH và FADH2. Các phân tử này sau đó sẽ vận chuyển những electron đó đến chuỗi chuyền điện tử, nơi phần lớn ATP sẽ được tạo ra.
Trước khi chu trình có thể bắt đầu, Pyruvate (sản phẩm 3-carbon của quá trình đường phân) phải được chuyển đổi thành Acetyl-CoA (một phân tử 2-carbon). Quá trình này được gọi là oxy hóa Pyruvate: một nhóm carboxyl (-COO⁻) được loại bỏ khỏi Pyruvate, giải phóng một phân tử carbon dioxide (CO2). Phân tử 2-carbon còn lại bị oxy hóa, và các electron mất đi được NAD+ thu nhận để tạo thành NADH. Phân tử 2-carbon đã bị oxy hóa (nhóm acetyl) được gắn vào Coenzyme A (CoA) để tạo thành Acetyl-CoA. Acetyl-CoA chính là phân tử nhiên liệu đầu vào cho chu trình Krebs.
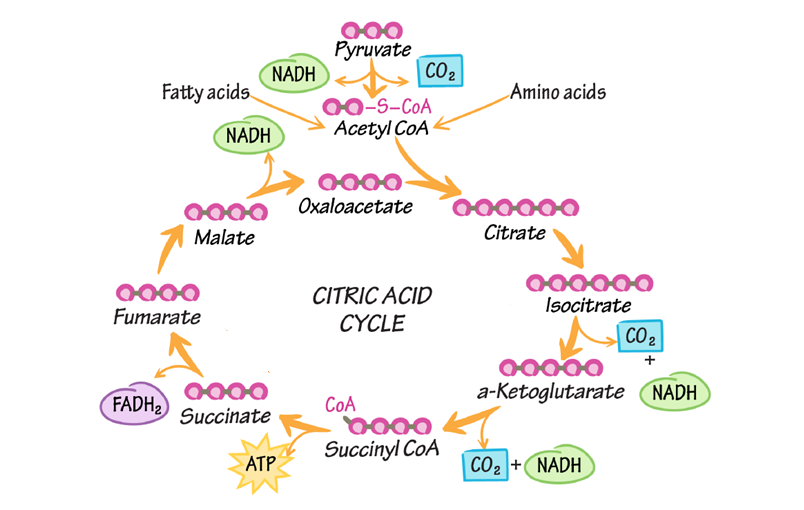
Chu trình axít citric là một vòng lặp gồm 8 bước, có nghĩa là phân tử cuối cùng của chu trình cũng là phân tử đầu tiên để bắt đầu một vòng mới. Mỗi vòng chu trình xử lý một phân tử Acetyl-CoA.
Mỗi vòng của chu trình Krebs tạo ra: 2 phân tử CO2 + 3 phân tử NADH + 1 phân tử FADH2 + 1 phân tử ATP (hoặc GTP). Vì mỗi phân tử glucose tạo ra hai Acetyl-CoA, tổng sản phẩm từ một phân tử glucose ban đầu là gấp đôi. Năng lượng thực sự khổng lồ được chứa trong 6 phân tử NADH và 2 phân tử FADH2 sẽ được rút ra ở giai đoạn tiếp theo.
Chuỗi truyền điện tử và Phosphoryl hóa oxy hóa
Đây là giai đoạn cuối cùng và cũng là giai đoạn tạo ra nhiều năng lượng nhất trong quá trình hô hấp hiếu khí.
Chuỗi chuyền điện tử (ETC) là một tập hợp các phân tử, chủ yếu là protein, được nhúng vào màng trong của ty thể. Màng này gấp nếp tạo thành các mào (cristae), làm tăng diện tích bề mặt để chứa hàng ngàn chuỗi chuyền điện tử. Các thành phần chính bao gồm:
- Bốn phức hợp protein lớn (I, II, III, IV) là các enzyme xuyên màng, đóng vai trò như các trạm trung chuyển electron.
- Hai phân tử mang electron di động là Ubiquinone (Coenzyme Q) - một phân tử lipid nhỏ, di chuyển tự do bên trong màng; Cytochrome c: - một protein nhỏ, di động ở phía không gian gian màng.
- ATP Synthase là một phức hợp enzyme khổng lồ, chịu trách nhiệm tổng hợp ATP.
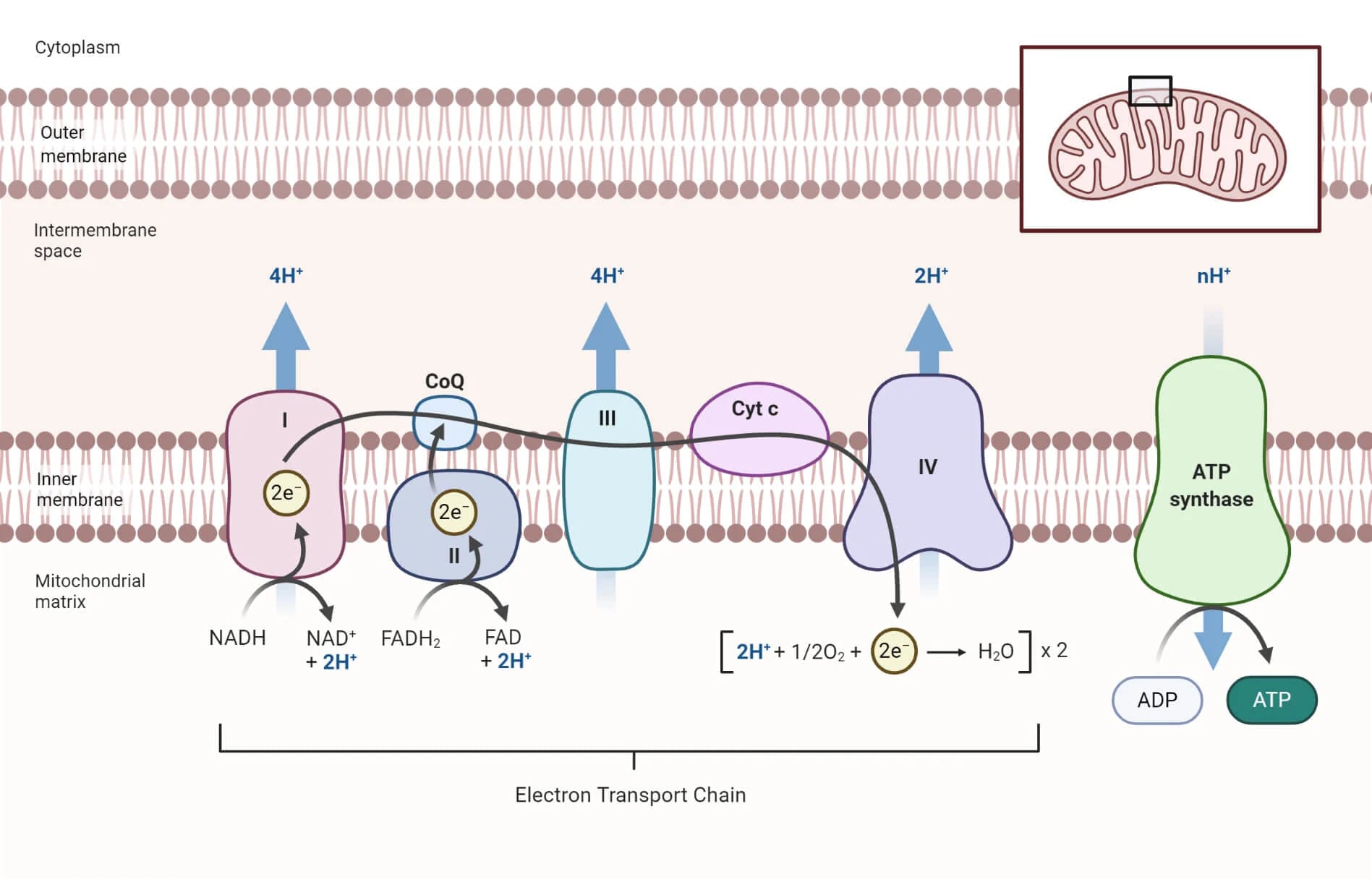
Dòng chảy của electron diễn ra theo một con đường xác định, từ mức năng lượng cao đến mức năng lượng thấp hơn, giống như một dòng thác:
- Tại Phức hợp I, NADH từ chu trình Krebs và đường phân nhường 2 electron năng lượng cao của nó, đồng thời chuyển trở lại thành NAD+. Năng lượng từ quá trình này được Phức hợp I sử dụng để bơm proton (H+) qua màng.
- Ở Phức hợp II, FADH2 chủ yếu từ chu trình Krebs nhường 2 electron của nó, chuyển lại thành FAD. Tuy nhiên, các electron này có mức năng lượng thấp hơn so với electron từ NADH, do đó Phức hợp II không bơm proton.
- Cả Phức hợp I và II đều chuyển electron của chúng cho phân tử mang di động Ubiquinone (Q). Q nhận electron, trở thành QH2, và di chuyển trong màng để đến Phức hợp III.
- QH2 nhường electron cho Phức hợp III. Năng lượng giải phóng ở đây tiếp tục được dùng để bơm thêm proton (H+) ra không gian gian màng. Sau đó, Phức hợp III chuyển các electron cho phân tử mang di động thứ hai là Cytochrome c.
- Cytochrome c mang electron đến Phức hợp IV, là trạm cuối cùng của dòng electron. Năng lượng được giải phóng một lần nữa để bơm proton (H+).
- Tại Phức hợp IV, các electron (đã ở mức năng lượng rất thấp) phải được loại bỏ khỏi chuỗi để chuỗi có thể tiếp tục hoạt động. Oxy chính là chất nhận electron cuối cùng. Một phân tử oxy (O2) sẽ nhận 4 electron và kết hợp với 4 proton (H+) từ chất nền để tạo thành 2 phân tử nước (H2O). Nếu không có oxy, chuỗi chuyền điện tử sẽ bị tắc nghẽn, NADH và FADH2 không thể được tái tạo, và toàn bộ quá trình sản xuất năng lượng sẽ ngừng lại.
Quá trình bơm proton liên tục đã tạo ra một nồng độ H+ rất cao ở không gian gian màng so với trong chất nền. Con đập năng lượng này (lực vận động proton) sau đó được khai thác để tổng hợp ATP. Các ion H+ không thể tự do khuếch tán qua màng trong ty thể. Chúng chỉ có một lối đi duy nhất để quay trở lại chất nền: đi xuyên qua kênh của enzyme ATP Synthase. Khi dòng proton chảy qua ATP synthase, nó làm cho một bộ phận của enzyme này quay giống như một tuabin. Năng lượng cơ học từ sự quay này được ATP synthase sử dụng để gắn một nhóm phosphate vô cơ (Pi) vào ADP (adenosine diphosphate), tạo ra ATP. Quá trình sử dụng năng lượng từ gradient hóa học (gradient proton) để thúc đẩy quá trình tổng hợp được gọi là hóa thẩm (chemiosmosis). Toàn bộ quá trình từ chuỗi chuyền điện tử đến hóa thẩm được gọi chung là phosphoryl hóa oxy hóa.
Mỗi phân tử NADH cung cấp đủ năng lượng để tổng hợp khoảng 2,5-3 phân tử ATP. Mỗi phân tử FADH2 tạo ra khoảng 1,5-2 phân tử ATP. Giai đoạn này tạo ra phần lớn ATP trong hô hấp tế bào, thu về khoảng 32-34 phân tử ATP, vượt xa so với sản lượng của đường phân và chu trình Krebs cộng lại.
Các con đường hô hấp đặc thù
Thực vật, với lối sống cố định, đã tiến hóa các cơ chế trao đổi chất linh hoạt để đối phó với những biến động của môi trường. Hô hấp ở thực vật không chỉ giới hạn ở con đường hiếu khí trung tâm mà còn bao gồm các con đường thay thế và các quá trình đặc thù khác.
Quang hô hấp
Quang hô hấp, đã nhắc đến ở bài viết Quang hợp, là một quá trình đặc biệt xảy ra ở thực vật C3 (như lúa, lúa mì, đậu nành) trong điều kiện cường độ ánh sáng và nhiệt độ cao, khi nồng độ CO2 trong lá giảm và nồng độ O2 tăng cao. Trong điều kiện này, enzyme RuBisCO, vốn có chức năng cố định CO2 trong quang hợp, lại hoạt động như một oxygenase, cố định CO2 vào phân tử RuBP. Phản ứng này tạo ra một hợp chất độc hại là phosphoglycolate, mà thực vật phải tiêu tốn năng lượng để tái chế thông qua một con đường phức tạp.
Quá trình này tiêu thụ O2 và giải phóng CO2 giống như hô hấp, nhưng không tạo ra ATP. Ngược lại, nó tiêu tốn năng lượng và làm mất đi các hợp chất carbon đã được cố định, do đó được coi là một quá trình lãng phí. RuBisCO là một enzyme cổ xưa, tiến hóa trong một môi trường có nồng độ CO2 cao và O2 thấp. Hoạt tính kép của nó là một di sản tiến hóa, trở nên bất lợi cho năng suất nông nghiệp trong điều kiện khí quyển hiện tại.
Con đường Oxidase thay thế (AOX)
Chuỗi chuyền điện tử trong ty thể thực vật có một đặc điểm độc đáo là sự phân nhánh. Bên cạnh con đường cytochrome chuẩn, còn có một con đường oxidase thay thế (AOX). Con đường AOX cho phép các electron từ bể ubiquinone được chuyển trực tiếp đến oxy để tạo thành nước (xúc tác bởi enzyme Alternative Oxidase), bỏ qua các phức hợp III và IV của chuỗi cytochrome. Vì bỏ qua hai vị trí bơm proton, con đường này không đi kèm với việc tổng hợp ATP. Năng lượng của các electron thay vì được tích trữ trong ATP sẽ được giải phóng dưới dạng nhiệt.
Con đường AOX không phải lúc nào cũng hoạt động. Nó được điều hòa chặt chẽ và chỉ được kích hoạt khi cần thiết. Enzyme AOX tồn tại ở dạng dimer không hoạt động (liên kết bởi cầu disulfide). Nó được kích hoạt khi các chất chuyển hóa như Pyruvate và các α-keto acid khác tích tụ, báo hiệu tình trạng dư thừa carbon, làm giảm các cầu disulfide và kích hoạt enzyme.
Một đặc tính quan trọng của AOX là nó không bị ức chế bởi các chất độc như cyanide, vốn tác động và làm tê liệt phức hợp IV của con đường cytochrome. Điều này cho phép hô hấp vẫn tiếp tục diễn ra ngay cả khi con đường chính bị ngộ độc.
Tại sao thực vật lại cần một con đường lãng phí năng lượng như vậy? AOX mang lại sự linh hoạt về mặt trao đổi chất và là một cơ chế thích nghi quan trọng.
- Tạo nhiệt (Thermogenesis): Đây là chức năng nổi tiếng nhất. Ở một số loài thực vật, như hoa của cây họ Ráy (Araceae), con đường AOX hoạt động mạnh mẽ để tạo ra một lượng nhiệt đáng kể. Nhiệt độ của hoa có thể cao hơn môi trường xung quanh tới 10-15°C. Lượng nhiệt này giúp bay hơi các hợp chất hữu cơ, thu hút côn trùng thụ phấn từ xa.
- Chống chịu stress Oxy hóa: Khi cây bị stress (ví dụ: cường độ ánh sáng quá cao, lạnh, hạn hán, mặn), chuỗi chuyền điện tử truyền thống có thể bị tắc nghẽn hoặc quá tải. Điều này dẫn đến việc các electron bị rò rỉ và phản ứng với oxy tạo ra các gốc Oxy hóa Phản ứng (Reactive Oxygen Species - ROS), gây hại cho tế bào. Con đường AOX hoạt động như một van an toàn, giúp giải tỏa áp lực electron, duy trì dòng chảy và giảm thiểu sự hình thành ROS.
- Cân bằng trao đổi chất: Khi cây quang hợp mạnh, lượng đường và các chất khử (NADH) được tạo ra có thể vượt quá nhu cầu ATP của tế bào. Nếu chuỗi chuyền điện tử truyền thống chậm lại do nồng độ ATP cao (cơ chế điều hòa ngược), các sản phẩm của quá trình đường phân và chu trình Krebs sẽ tích tụ, ức chế ngược lại chính các con đường này. AOX cho phép quá trình oxy hóa NADH và FADH₂ tiếp tục diễn ra mà không cần sản xuất ATP, giúp duy trì hoạt động của chu trình Krebs và đường phân, đảm bảo cung cấp các khung carbon cần thiết cho các quá trình sinh tổng hợp khác.
Hô hấp kỵ khí
Khi thực vật bị thiếu oxy, chúng không thể thực hiện chuỗi chuyền điện tử và chu trình Krebs. Thay vào đó, chúng chuyển sang một quá trình thay thế gọi là hô hấp kỵ khí hay phổ biến hơn là lên men. Đây là một cơ chế sinh tồn ngắn hạn, giúp tế bào thực vật tạo ra một lượng nhỏ năng lượng đủ để duy trì các hoạt động sống cơ bản cho đến khi có lại oxy.
Hô hấp kỵ khí xảy ra khi các tế bào hoặc mô thực vật không được cung cấp đủ oxy. Rễ cây bị úng nước là trường hợp phổ biến nhất. Đất bị ngập nước sẽ đẩy hết không khí ra ngoài, khiến rễ cây rơi vào tình trạng thiếu oxy nghiêm trọng.
Giai đoạn đường phân hoàn toàn giống với giai đoạn đường phân trong hô hấp hiếu khí. Trong điều kiện thiếu oxy tạm thời, Pyruvate được khử trực tiếp bởi NADH để tạo thành lactate (axit lactic). Phản ứng này được xúc tác bởi enzyme Lactate dehydrogenase. Tuy nhiên, việc tích tụ lactate sẽ làm giảm độ pH của tế bào chất (gây nhiễm toan), ức chế hoạt động của nhiều enzyme và gây hại cho tế bào. Do đó, con đường này chỉ diễn ra trong một thời gian ngắn.
Khi tình trạng thiếu oxy kéo dài và pH bắt đầu giảm, thực vật sẽ chuyển sang con đường lên men etylic. Đây là con đường chính trong hô hấp kỵ khí của thực vật. Pyruvate bị loại bỏ một phân tử CO2 (khử carboxyl) để tạo thành Acetaldehyde. Phản ứng được xúc tác bởi enzyme Pyruvate decarboxylase. Acetaldehyde sau đó bị khử bởi NADH để tạo thành ethanol. Phản ứng này tái tạo lại NAD+ và được xúc tác bởi enzyme Alcohol dehydrogenase. Con đường này ưu việt hơn lên men lactic vì ethanol là một chất trung tính, không làm thay đổi pH của tế bào. Tuy nhiên, nếu tích tụ ở nồng độ cao trong thời gian dài, ethanol cũng trở thành một chất độc gây chết tế bào.
Toàn bộ quá trình hô hấp kỵ khí chỉ tạo ra 2 ATP từ một phân tử glucose (so với khoảng 36-38 ATP của hô hấp hiếu khí). Năng lượng còn lại phần lớn vẫn bị khóa trong các phân tử ethanol.
Các yếu tố điều chỉnh cường độ hô hấp
Cường độ hô hấp của thực vật không phải là một hằng số mà thay đổi mạnh mẽ tùy thuộc vào các yếu tố bên trong và điều kiện môi trường bên ngoài.
Cường độ hô hấp không đồng đều trên khắp cơ thể thực vật. Nó diễn ra mạnh nhất ở các mô có hoạt động trao đổi chất cao như hạt đang nảy mầm, mô phân sinh, hoa đang phát triển, và quả non. Ngược lại, các mô ở trạng thái ngủ nghỉ như hạt khô có cường độ hô hấp rất thấp.
Trong một giới hạn nhất định, cường độ hô hấp tăng theo nhiệt độ, đạt tối ưu ở khoảng 30-35°C. Nhiệt độ cao hơn mức này sẽ làm biến tính các enzyme hô hấp, khiến cường độ giảm mạnh.
Nước là dung môi thiết yếu cho các phản ứng enzyme. Cường độ hô hấp tỷ lệ thuận với hàm lượng nước trong mô. Tình trạng thiếu nước làm giảm hô hấp, trong khi việc cung cấp nước cho hạt khô sẽ làm cường độ hô hấp tăng vọt, khởi động quá trình nảy mầm.
Oxy (O2) là nguyên liệu trực tiếp cho hô hấp hiếu khí, nồng độ O2 ảnh hưởng trực tiếp đến cường độ hô hấp. Trong điều kiện thiếu oxy, chẳng hạn như ở đất bị ngập úng, hô hấp hiếu khí bị ức chế, buộc tế bào phải chuyển sang con đường lên men kỵ khí kém hiệu quả hơn nhiều.
Carbon dioxide (CO2) là một sản phẩm của hô hấp, nồng độ CO2 cao trong môi trường có thể gây ra hiện tượng ức chế ngược, làm chậm quá trình hô hấp. Nguyên tắc này là nền tảng cho nhiều kỹ thuật bảo quản nông sản.
Hô hấp là một phần của một mạng lưới trao đổi chất rộng lớn, có mối liên hệ mật thiết với quang hợp, có những ứng dụng quan trọng trong nông nghiệp và công nghệ thực phẩm.


