
Quang hợp
Pha sáng, Calvin, C3, C4, CAM
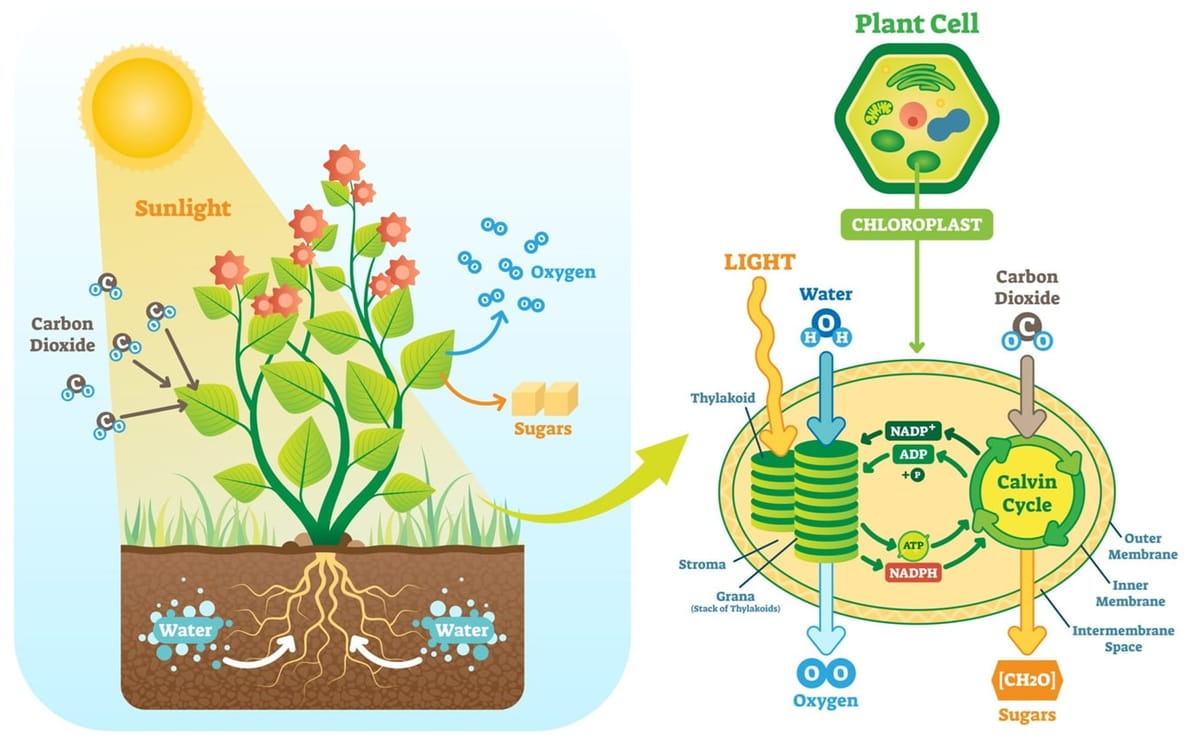
Quang hợp là quá trình sinh hóa quan trọng bậc nhất trên Trái Đất, tạo nền tảng cho gần như toàn bộ chuỗi thức ăn và chi phối các chu trình carbon và oxy toàn cầu. Quá trình này, trong đó năng lượng ánh sáng được chuyển đổi thành năng lượng hóa học dưới dạng các hợp chất hữu cơ, diễn ra bên trong một bào quan chuyên biệt cao độ gọi là lục lạp.
Cấu tạo lục lạp
Lục lạp, thường có hình bầu dục, hình đĩa hoặc hình cầu, là bào quan thực hiện chức năng quang hợp và chỉ có ở tế bào thực vật và một số sinh vật nguyên sinh. Kích thước và số lượng của chúng thay đổi tùy thuộc vào loại tế bào và mô, ví dụ, lục lạp trong tế bào mô giậu thường lớn hơn và nhiều hơn so với lục lạp trong tế bào bao bó mạch.
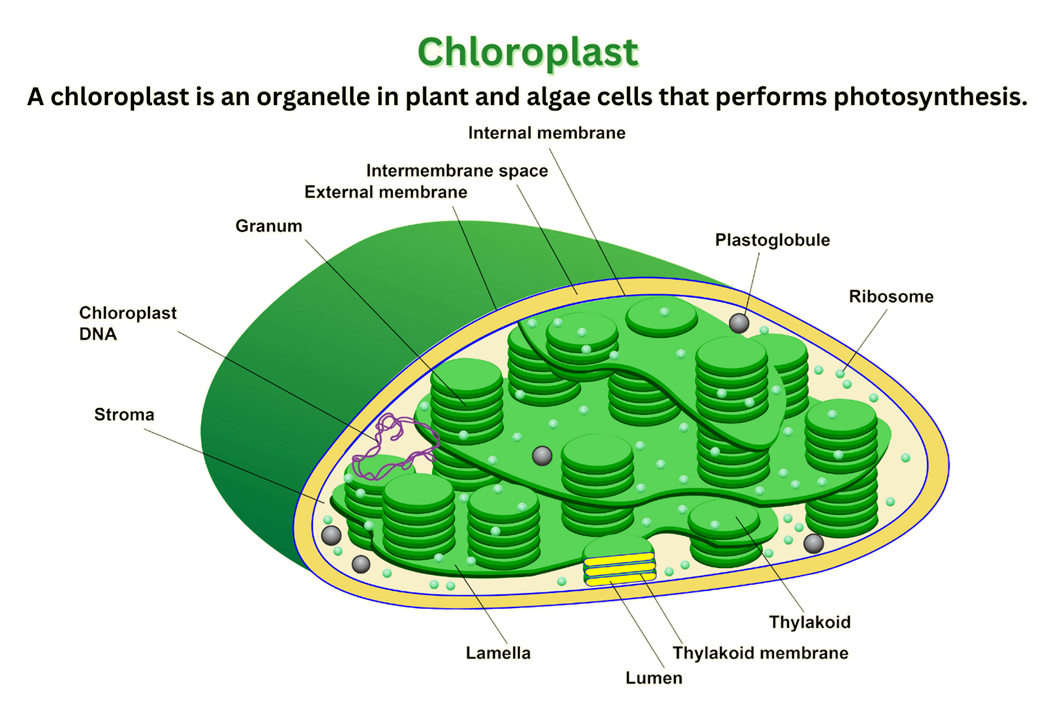
Lục lạp được bao bọc bởi một lớp màng kép lipoprotein, gồm màng ngoài và màng trong. Màng ngoài có tính thấm cao, chứa nhiều protein vận chuyển lớn gọi là porin, tạo điều kiện thuận lợi cho việc trao đổi các chất chuyển hóa giữa lục lạp và tế bào chất. Màng trong có tính thấm chọn lọc hơn nhiều, chứa các protein vận chuyển chuyên biệt để kiểm soát chặt chẽ dòng chảy của các ion và các sản phẩm của quang hợp, chẳng hạn như đường và ATP. Cả hai màng đều trơn nhẵn, không gấp nếp như màng trong của ty thể. Khoang gian màng là không gian hẹp nằm giữa hai lớp màng. Hệ thống màng kép này không chỉ là một rào cản vật lý mà còn là bằng chứng quan trọng củng cố cho thuyết nội cộng sinh về nguồn gốc của lục lạp.
Bên trong màng trong là một dung dịch dạng lỏng, giàu protein được gọi là chất nền stroma. Đây là nơi diễn ra các phản ứng của pha tối, hay còn gọi là chu trình Calvin. Stroma chứa một hệ thống enzyme dày đặc, nổi bật nhất là RuBisCO, enzyme xúc tác cho quá trình cố định CO2. Ngoài ra, stroma còn chứa hệ thống di truyền riêng của lục lạp, bao gồm DNA dạng vòng và các ribosome 70S, cùng với các hạt tinh bột là nơi dự trữ năng lượng tạm thời.
Nằm lơ lửng trong stroma là một mạng lưới nội màng phức tạp gồm các túi dẹt thông nhau gọi là thylakoid. Đây chính là nơi diễn ra các phản ứng phụ thuộc ánh sáng (pha sáng) của quang hợp. Các thylakoid thường xếp chồng lên nhau tạo thành các cấu trúc gọi là granum (số nhiều: grana). Cấu trúc xếp chồng này làm tăng đáng kể tỷ lệ diện tích bề mặt trên thể tích, tối đa hóa khả năng hấp thụ ánh sáng và hiệu quả của chuỗi chuyền electron. Lamella là các thylakoid không xếp chồng, có chức năng nối liền các grana với nhau, tạo thành một hệ thống màng liên tục và một khoang duy nhất bên trong gọi là xoang thylakoid (thylakoid lumen). Sự sắp xếp này đảm bảo toàn bộ hệ thống hoạt động như một thể thống nhất.
Sự phân chia không gian bên trong lục lạp là một kiệt tác của kỹ thuật sinh học. Việc tách biệt pha sáng (diễn ra trên màng thylakoid) và chu trình Calvin (diễn ra trong stroma) cho phép điều hòa độc lập hai quá trình này và duy trì các môi trường hóa học riêng biệt. Cụ thể, chuỗi chuyền electron trên màng thylakoid bơm các proton (H+) từ stroma vào xoang thylakoid, tạo ra một gradient điện hóa dốc. Các proton này chỉ có thể quay trở lại stroma thông qua phức hệ ATP synthase, một bộ máy phân tử khai thác dòng chảy proton để tổng hợp ATP. Nếu hai pha này không được ngăn cách, gradient proton thiết yếu này không thể được thiết lập và duy trì một cách hiệu quả, làm tê liệt quá trình sản xuất ATP. Do đó, sự phân ngăn này không phải là ngẫu nhiên mà là yếu tố cốt lõi cho hiệu suất của toàn bộ quá trình quang hợp.
Cấu trúc lục lạp cũng thể hiện sự thích nghi với môi trường. Ví dụ, thực vật C4 sở hữu hai loại lục lạp khác nhau về cấu trúc và chức năng: lục lạp ở tế bào mô giậu và lục lạp ở tế bào bao bó mạch, một sự thích nghi chuyên biệt cho điều kiện khô hạn và nồng độ CO2 thấp.
Lục lạp sở hữu hệ thống di truyền riêng, được gọi là DNA lục lạp (cpDNA). Đây thường là một phân tử DNA mạch vòng, kép, có chiều dài khoảng 120.000-170.000 cặp base. Bộ gen này mã hóa cho một số protein và RNA cần thiết cho chức năng của chính lục lạp. Một đặc điểm đáng chú ý của nhiều cpDNA là sự hiện diện của hai vùng lặp lại đảo ngược (inverted repeats - IRs), giúp ổn định cấu trúc của bộ gen.
Stroma chứa các ribosome 70S, có cấu trúc tương tự ribosome của sinh vật nhân sơ (vi khuẩn) và khác biệt với ribosome 80S trong tế bào chất của tế bào thực vật nhân thực. Các ribosome này chịu trách nhiệm dịch mã các mRNA được phiên mã từ cpDNA để tổng hợp protein ngay tại lục lạp.
Sự tồn tại của một bộ gen riêng và các ribosome kiểu nhân sơ là bằng chứng vững chắc cho thuyết nội cộng sinh, cho rằng lục lạp có nguồn gốc từ một vi khuẩn lam quang hợp tự do đã bị một tế bào nhân thực sơ khai nuốt chửng và hình thành mối quan hệ cộng sinh. Điều này giải thích tại sao lục lạp có khả năng tự nhân đôi và tổng hợp một phần protein cho riêng mình, nhưng vẫn phụ thuộc phần lớn vào các gen trong nhân tế bào. Hơn nữa, cpDNA thường được di truyền theo dòng mẹ, một khái niệm then chốt trong di truyền và chọn giống thực vật.
Sắc tố quang hợp
Các sắc tố quang hợp được sắp xếp thành các phức hệ thu nhận ánh sáng nằm trên màng thylakoid, có chức năng hấp thụ và truyền năng lượng ánh sáng.
Diệp lục (Chlorophylls) là nhóm sắc tố chính, tạo nên màu xanh lục đặc trưng của thực vật.
- Diệp lục a (C55H72O5N4Mg): Là sắc tố quang hợp chính, có mặt trong trung tâm phản ứng của các hệ quang hợp (photosystems). Nó hấp thụ mạnh nhất ở vùng ánh sáng xanh tím (khoảng 430 nm) và đỏ (khoảng 662 nm) và trực tiếp tham gia vào quá trình chuyển đổi năng lượng.
- Diệp lục b (C55H72O6N4Mg): Là sắc tố phụ, hấp thụ ánh sáng ở các bước sóng hơi khác so với diệp lục a và truyền năng lượng đã hấp thụ cho diệp lục a, qua đó mở rộng phổ ánh sáng hữu dụng cho quang hợp.
Carotenoids là nhóm sắc tố phụ có màu đỏ, cam hoặc vàng, bao gồm carotene và xanthophyll. Chúng hấp thụ năng lượng ở vùng ánh sáng xanh-lục, nơi diệp lục hấp thụ kém, và chuyển năng lượng này đến trung tâm phản ứng. Dưới cường độ ánh sáng mạnh, carotenoids có vai trò quan trọng trong việc tản nhiệt năng lượng ánh sáng dư thừa, bảo vệ các phân tử diệp lục khỏi bị tổn thương do oxy hóa quang học (hiện tượng "cháy nắng").
Một hệ thống đa sắc tố là sự thích nghi để tối đa hóa việc thu nhận năng lượng và đảm bảo sự sống còn. Một loài thực vật chỉ dựa vào diệp lục a sẽ hoạt động kém hiệu quả vì nó sẽ "mù" trước phần lớn phổ ánh sáng nhìn thấy. Sự tiến hóa của các sắc tố phụ là một chiến lược để lấp đầy những khoảng trống, cho phép thực vật tận dụng nhiều hơn ánh sáng mặt trời sẵn có. Vai trò kép của carotenoids vừa là bộ thu ánh sáng vừa là chất bảo vệ quang học thể hiện một giải pháp tiến hóa tinh vi cho vấn đề tối ưu hóa việc thu nhận năng lượng trong khi giảm thiểu thiệt hại do chính năng lượng đó gây ra. Thành phần sắc tố của một chiếc lá là sự cân bằng được tinh chỉnh giữa việc tối đa hóa thu nhận ánh sáng và ngăn chặn sự tự hủy hoại, một sự cân bằng thay đổi tùy thuộc vào môi trường ánh sáng của cây (ví dụ: lá cây ưa sáng và lá cây ưa bóng).
Phản ứng pha sáng
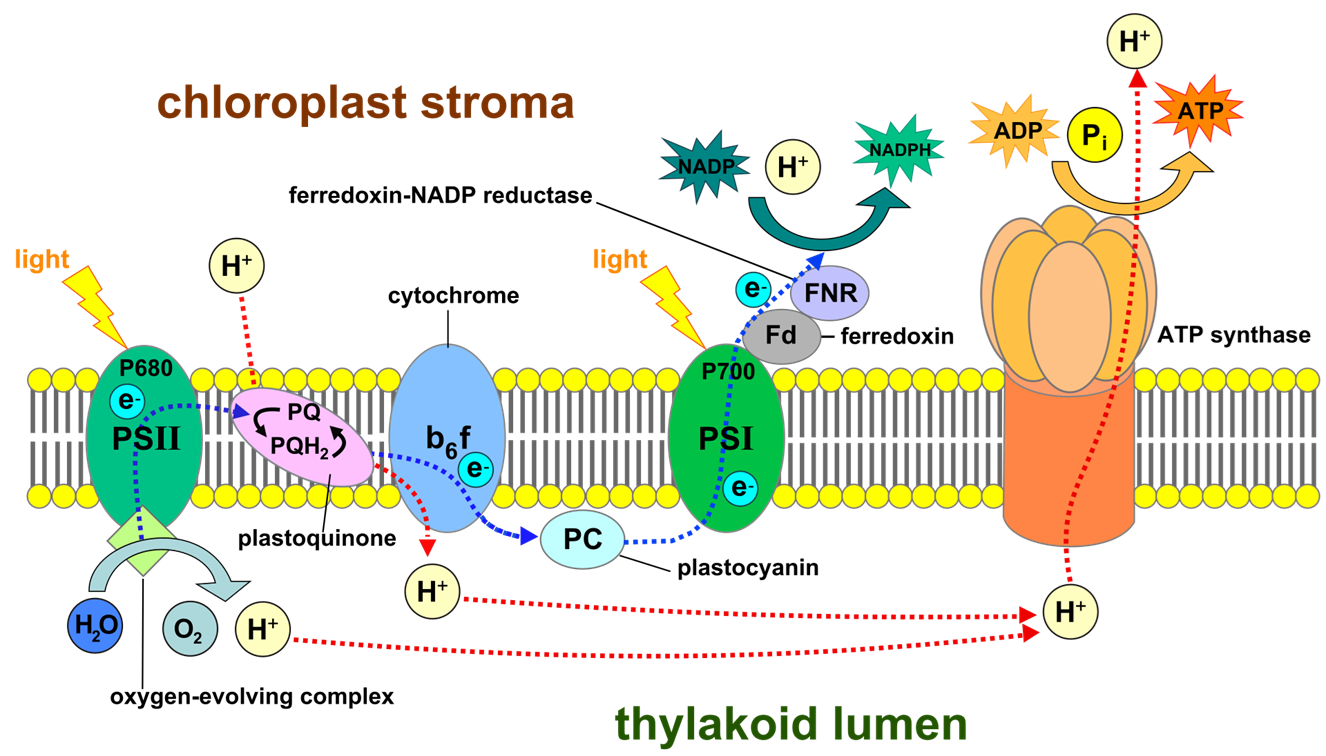
Pha sáng, hay còn gọi là phản ứng phụ thuộc vào ánh sáng, là giai đoạn đầu tiên của quá trình quang hợp. Giai đoạn này chuyển hóa năng lượng ánh sáng thành năng lượng hóa học dưới dạng các phân tử mang năng lượng là ATP và NADPH. Pha sáng diễn ra tại màng thylakoid bên trong lục lạp.
Diệp lục và các sắc tố quang hợp khác trong màng thylakoid hấp thụ năng lượng từ ánh sáng mặt trời. Năng lượng này kích thích các electron trong phân tử diệp lục, đưa chúng lên mức năng lượng cao hơn. Một photon ánh sáng đập vào một phân tử sắc tố trong Hệ quang hợp II (PSII), năng lượng được truyền từ phân tử này sang phân tử khác cho đến khi đến trung tâm phản ứng P680.
Phân tử P680 bị kích thích sẽ nhường một electron cho một chất nhận electron sơ cấp. P680 lúc này trở thành một chất oxy hóa cực mạnh (P680+) và ngay lập tức oxy hóa một phân tử nước, tách lấy các electron để bù lại electron đã mất. Quá trình này giải phóng khí oxy (O2) và các ion hydro (H+) vào xoang thylakoid (quang phân ly nước). Phương trình tổng quát là 2H2O → 4H+ + 4e− + O2.
Các electron bị kích thích từ PSII được truyền đến PSI thông qua một chuỗi chuyền electron. Sự rơi của các electron xuống mức năng lượng thấp hơn được sử dụng để bơm H+ từ stroma qua màng vào xoang thylakoid, tạo ra một lực vận động proton.
Gradient nồng độ H+ cao trong xoang thylakoid cung cấp năng lượng cho enzyme ATP synthase hoạt động, xúc tác cho quá trình phosphoryl hóa ADP thành ATP. Quá trình này được gọi là quang phosphoryl hóa.
Năng lượng ánh sáng cũng kích thích trung tâm phản ứng P700 của PSI, khiến nó nhường electron cho một chuỗi chuyền electron thứ hai. Các electron này cuối cùng được chuyển đến NADP+, khử nó thành NADPH.
Phương trình tổng quát cho các phản ứng phụ thuộc vào ánh sáng là:
12H2O + 12NADP+ + 18ADP + 18Pi + Năng lượng ánh sáng → 6O2 + 12NADPH + 18ATP
Tóm lại, pha sáng của quang hợp sử dụng năng lượng ánh sáng và nước để tạo ra ATP và NADPH. Hai phân tử mang năng lượng này sau đó sẽ được sử dụng trong pha tối (chu trình Calvin) để tổng hợp đường và các hợp chất hữu cơ khác từ CO2.
Chu trình Calvin
Chu trình Calvin, hay còn gọi là pha tối hoặc phản ứng không phụ thuộc vào ánh sáng, là giai đoạn thứ hai của quá trình quang hợp, được đặt theo tên của nhà khoa học Melvin Calvin, người đã khám phá ra nó. Giai đoạn này diễn ra trong chất nền (stroma) của lục lạp. Mục đích chính của chu trình Calvin là sử dụng năng lượng hóa học (ATP và NADPH) được tạo ra từ pha sáng để chuyển hóa carbon dioxide (CO2) thành đường (glucose) và các hợp chất hữu cơ khác. Mặc dù được gọi là "pha tối", nó không nhất thiết phải diễn ra trong bóng tối mà chỉ đơn giản là không trực tiếp cần ánh sáng, nhưng nó hoàn toàn phụ thuộc vào các sản phẩm của pha sáng.
Một phân tử CO2 từ không khí được kết hợp với một phân tử hữu cơ 5-carbon có sẵn trong lục lạp là ribulose-1,5-bisphosphate (RuBP). Phản ứng này được xúc tác bởi enzyme RuBisCO. Sản phẩm tạo thành là một hợp chất 6-carbon không bền, ngay lập tức bị tách ra thành hai phân tử 3-phosphoglycerate (3-PGA), là các hợp chất 3-carbon.
Năng lượng từ ATP và các electron từ NADPH (cả hai đều là sản phẩm của pha sáng) được sử dụng để chuyển đổi mỗi phân tử 3-PGA thành một phân tử đường 3-carbon khác là glyceraldehyde-3-phosphate (G3P). Trong quá trình này, ATP bị biến đổi thành ADP và NADPH bị oxy hóa thành NADP+. ADP và NADP+ sau đó quay trở lại pha sáng để được tái tạo.
Trong sáu phân tử G3P được tạo ra, chỉ có một phân tử được coi là sản phẩm ròng của chu trình và sẽ rời khỏi chu trình để tế bào thực vật tổng hợp nên glucose, cellulose, tinh bột và các hợp chất hữu cơ khác. Năm phân tử G3P còn lại sẽ ở lại chu trình. Chúng sử dụng năng lượng từ ATP để tái sắp xếp và tái tạo lại ba phân tử RuBP ban đầu. Khi RuBP được tái tạo, chu trình lại sẵn sàng để cố định thêm CO2 và tiếp tục.
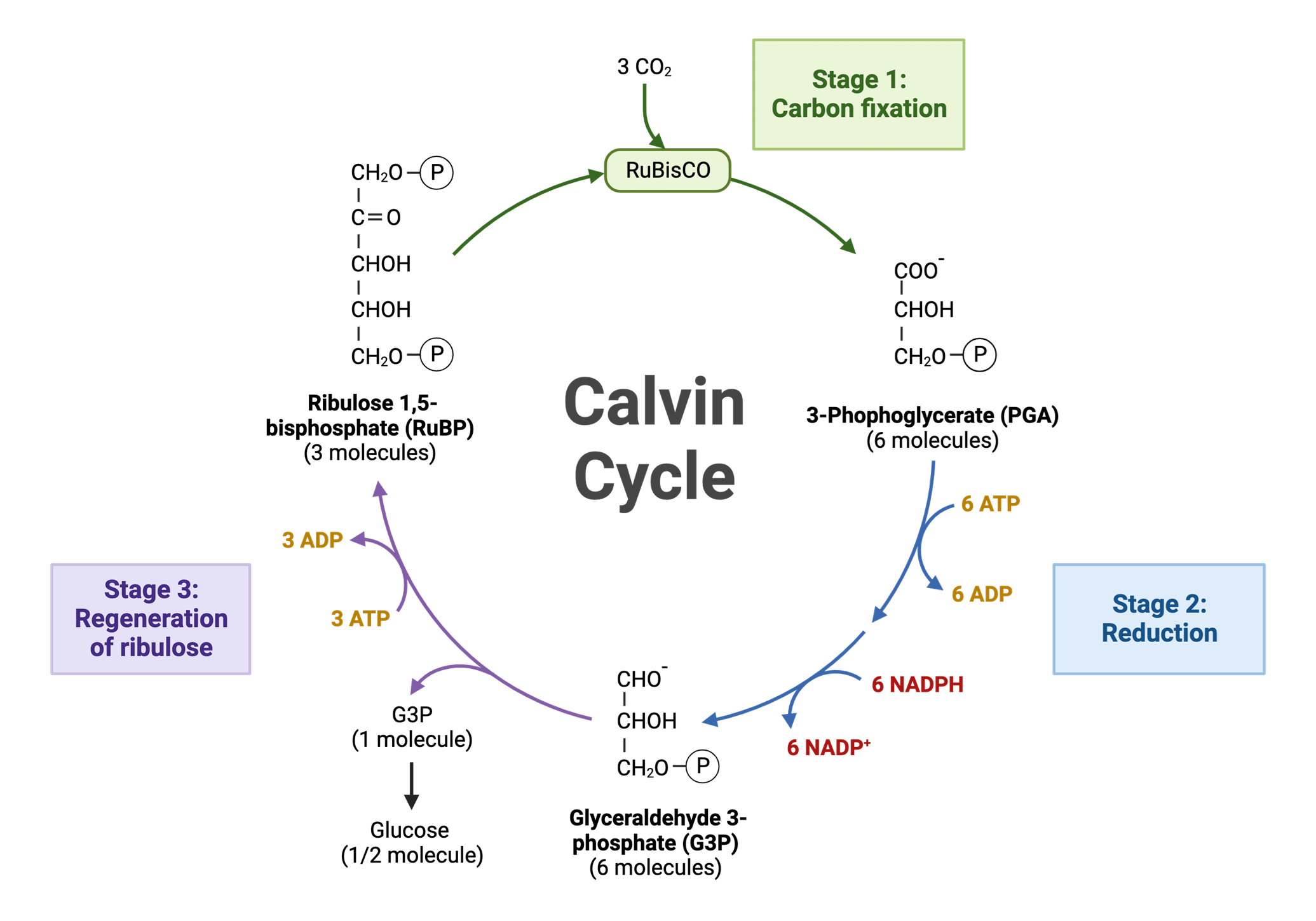
Để tạo ra một phân tử G3P (sản phẩm ròng), chu trình phải diễn ra 3 lần, cố định 3 phân tử CO2. Hai phân tử G3P sau đó có thể kết hợp với nhau để tạo ra một phân tử glucose (6-carbon), nguồn năng lượng chính cho thực vật và hầu hết các sinh vật sống trên Trái Đất.
Tổng hợp cả hai pha, phương trình quang hợp thường được viết:
6CO2 + 12H2O + [quang năng] → C6H12O6 + 6O2 + 6H2O
Pha sáng và chu trình Calvin được liên kết chặt chẽ về mặt sinh hóa; một pha không thể diễn ra bền vững nếu thiếu pha kia. ATP và NADPH được tạo ra ở thylakoid là "đồng tiền năng lượng" để "chi trả" cho việc cố định carbon trong stroma. Điều này tạo ra một vòng điều hòa chặt chẽ. Khi không có ánh sáng, việc sản xuất ATP và NADPH ngừng lại, và chu trình Calvin nhanh chóng dừng theo. Sự kết hợp này đảm bảo rằng thực vật không lãng phí năng lượng để cố định carbon khi không có năng lượng từ ánh sáng. Đây không chỉ là sự phụ thuộc một chiều. Chu trình Calvin tái tạo ADP và NADP+, là những cơ chất thiết yếu cho pha sáng. Nếu không có sự tái tạo này, pha sáng cũng sẽ ngừng lại. Điều này tạo ra một chu trình tự điều chỉnh hoàn hảo, nơi tốc độ của một quá trình ảnh hưởng trực tiếp đến tốc độ của quá trình kia, đảm bảo sự cân bằng trao đổi chất.
Một điểm đáng chú ý là chức năng kép của enzyme RuBisCO, đây là một nút thắt cổ chai lớn trong tiến hóa. RuBisCO có thể liên kết với O2 cũng như CO2. Khi liên kết với O2, nó khởi đầu một quá trình lãng phí gọi là quang hô hấp (photorespiration), tiêu thụ ATP và giải phóng CO2 đã được cố định trước đó, làm giảm hiệu suất quang hợp. Sự kém hiệu quả này trở nên trầm trọng hơn trong điều kiện nóng và khô, khi thực vật đóng khí khổng để tiết kiệm nước, làm cho nồng độ O2 tăng lên và nồng độ CO2 giảm xuống bên trong lá. RuBisCO đã tiến hóa trong một bầu khí quyển cổ đại có rất ít oxy và nhiều carbon dioxide. Trong môi trường đó, sự thiếu chuyên biệt của nó không phải là một vấn đề. Khi quá trình quang hợp của vi khuẩn lam và thực vật giải phóng một lượng lớn oxy vào khí quyển qua hàng tỷ năm, enzyme tồn tại từ trước này đột nhiên ở trong một môi trường mà khiếm khuyết của nó trở thành một gánh nặng đáng kể. Bối cảnh lịch sử này giải thích tại sao một enzyme quan trọng như vậy lại kém hiệu quả đến thế. Sự kém hiệu quả này là áp lực chọn lọc chính dẫn đến sự tiến hóa của quang hợp C4 và CAM, là những cơ chế phức tạp để cô đặc carbon đã tiến hóa độc lập nhiều lần nhằm khắc phục những hạn chế của RuBisCO.
Cây C3, C4, CAM
Để thích nghi với các điều kiện môi trường khác nhau, đặc biệt là khí hậu nóng và khô, thực vật đã phát triển các con đường quang hợp khác nhau. Phổ biến nhất là C3, nhưng C4 và CAM là những biến thể quan trọng giúp cây tồn tại và phát triển hiệu quả hơn trong điều kiện khắc nghiệt.
Cây C3 (con đường quang hợp tiêu chuẩn) đại diện cho hầu hết các loài thực vật trên Trái Đất, bao gồm lúa mì, lúa gạo, đậu nành, khoai tây, và các cây thân gỗ. Enzyme cố định CO2: RuBisCO. Sản phẩm ổn định đầu tiên là 3-phosphoglycerate (3-PGA), là lý do con đường này được gọi là "C3". Cấu trúc lá không có cấu trúc giải phẫu đặc biệt. Tế bào mô giậu và tế bào bao bó mạch có cấu trúc lục lạp giống nhau. Hiện tượng hô hấp sáng xảy ra mạnh và gây lãng phí.
Cây C4 (thích nghi với điều kiện nóng, ẩm) với đại diện điển hình là các loài cây nhiệt đới như mía, ngô (bắp), cao lương, và nhiều loài cỏ. Quá trình bắt đầu từ việc hình thành các mô tiền dẫn, sẽ phát triển thành gân lá. Các tế bào xung quanh mô tiền dẫn được phát triển thành hai loại riêng biệt:
- Tế bào bao bó mạch (Bundle Sheath - BS): Vòng tế bào bên trong, ngay sát bó mạch. Chúng phát triển lớn hơn, có thành tế bào dày (thường không thấm khí CO2), và chứa các lục lạp lớn, thường giàu tinh bột nhưng ít cấu trúc grana.
- Tế bào thịt lá, mô giậu (Mesophyll - M): Vòng tế bào bên ngoài, bao quanh lớp tế bào bao bó mạch. Chúng có thành mỏng hơn và chứa các lục lạp bình thường với hệ thống grana phát triển để thực hiện pha sáng hiệu quả.
Hai loại tế bào phát triển hai loại lục lạp khác nhau về mặt hình thái và chức năng để đảm nhận các phần riêng biệt của chu trình quang hợp C4. Mạng lưới các cầu sinh chất (plasmodesmata) dày đặc hình thành giữa tế bào M và tế bào BS để đảm bảo sự vận chuyển nhanh chóng các chất chuyển hóa (axit 4-carbon). Cấu trúc lá có cấu trúc giải phẫu đặc trưng gọi là giải phẫu Kranz. Các tế bào bao bó mạch phát triển lớn, có thành dày và chứa nhiều lục lạp, xếp sít sao quanh các bó mạch. Các tế bào thịt lá bao quanh bên ngoài. Sự phân chia không gian này là chìa khóa cho cơ chế C4.
Tại tế bào thịt lá (mô giậu, M), các gen mã hóa cho enzyme PEP carboxylase (PEPC) được biểu hiện mạnh mẽ. Enzyme này có ái lực cao với CO2 và thực hiện bước cố định CO2 đầu tiên, tạo ra hợp chất 4-carbon. Enzyme RuBisCO gần như không được biểu hiện ở loại tế bào này. Chất nhận CO2 đầu tiên là Phosphoenolpyruvate (PEP) sau khi kết hợp với CO2 tạo sản phẩm ổn định đầu tiên là Oxaloacetate (một hợp chất 4-carbon), là lý do con đường này được gọi là "C4". Chất này sau đó được chuyển thành malate hoặc aspartate.
Tại tế bào bao bó mạch, Các gen mã hóa cho enzyme RuBisCO và các enzyme khác của chu trình Calvin (chu trình C3) được biểu hiện ở mức độ cao. Các enzyme có chức năng giải phóng CO2 từ hợp chất 4-carbon (ví dụ: enzyme malic) cũng được tập trung tại đây. Điều này tạo ra một buồng nén CO2, làm tăng nồng độ CO2 xung quanh RuBisCO lên gấp nhiều lần so với không khí, giúp loại bỏ hiện tượng hô hấp sáng. Hiệu suất quang hợp cao hơn trong điều kiện cường độ ánh sáng và nhiệt độ cao, sử dụng nước hiệu quả hơn.
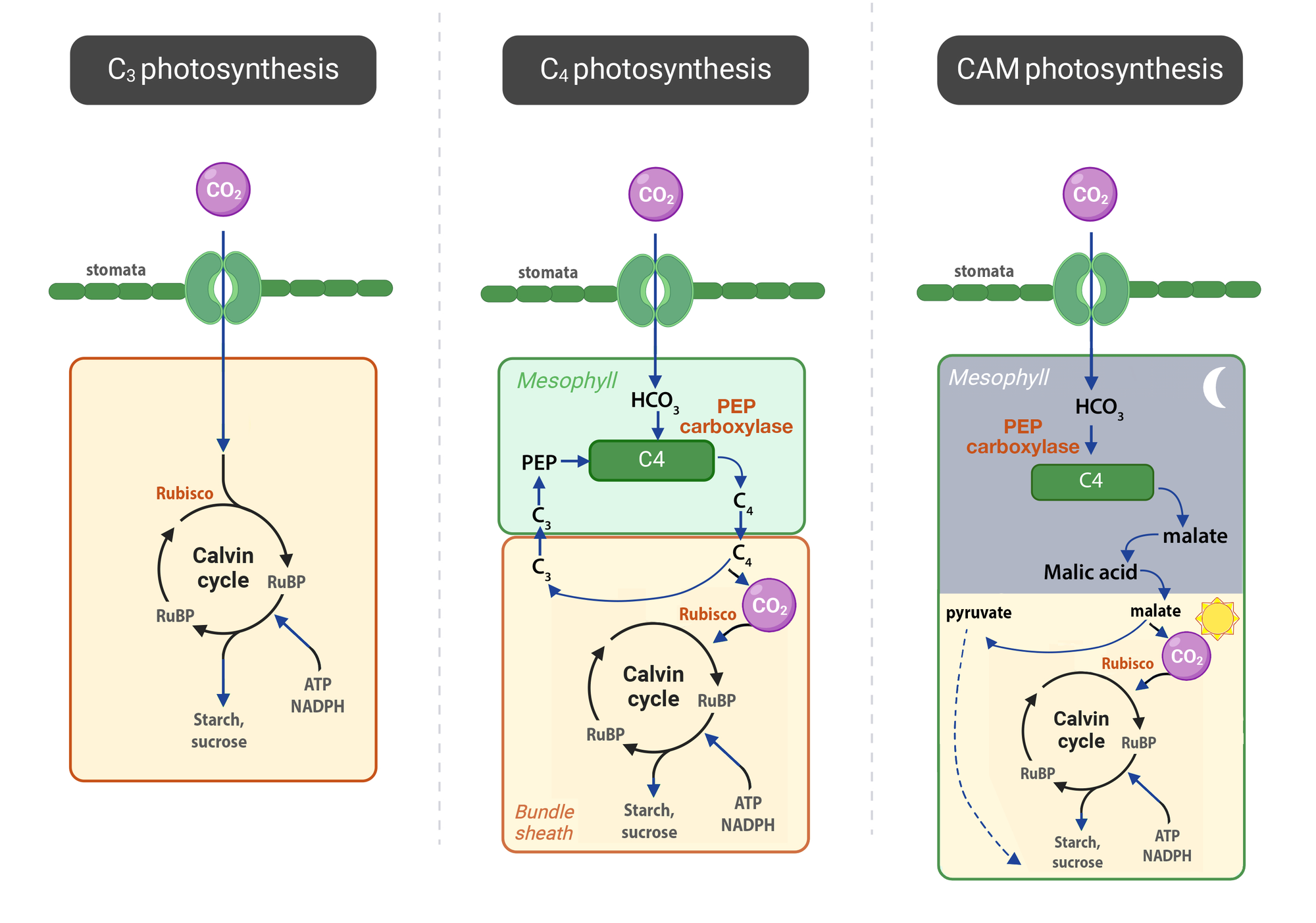
Quá trình phát triển của thực vật CAM (Crassulacean Acid Metabolism) là một câu chuyện hấp dẫn về sự thích nghi của thực vật với những môi trường khắc nghiệt nhất trên Trái Đất, như sa mạc và vùng khô hạn. Không giống như cây C4 phát triển cấu trúc giải phẫu chuyên biệt, sự phát triển của cây CAM chủ yếu tập trung vào những thay đổi về sinh lý và giải phẫu tế bào để tối ưu hóa việc sử dụng nước. Các tế bào thịt lá (mesophyll) phát triển với kích thước rất lớn, thành tế bào mỏng cho phép chúng phình to để chứa nước. Trung tâm của tế bào CAM là một không bào khổng lồ. Trong quá trình phát triển, không bào này trở thành nơi dự trữ axit malic được tạo ra vào ban đêm. Kích thước lớn của nó là tối quan trọng cho khả năng quang hợp CAM. Bề mặt lá hoặc thân cây phát triển một lớp biểu bì dày, được bao phủ bởi một lớp sáp không thấm nước. Cấu trúc này hoạt động như một lớp rào cản, ngăn chặn sự bay hơi nước từ bề mặt cây vào ban ngày khi trời nóng.
Sự phát triển của cây CAM không phải là thay đổi cấu trúc lá thành hai loại tế bào riêng biệt như cây C4, mà là thiết lập một nhịp điệu sinh học ngày-đêm chặt chẽ bên trong cùng một loại tế bào (tế bào thịt lá).
Ban đêm, enzyme PEP carboxylase (PEPC) được kích hoạt mạnh mẽ, cố định CO2 lấy từ không khí (khi khí khổng mở) thành axit oxaloacetic, sau đó nhanh chóng chuyển thành axit malic. Các kênh vận chuyển trên màng không bào được kích hoạt để bơm axit malic vào bên trong không bào. Nồng độ axit trong không bào tăng dần suốt đêm, làm cho dịch bào trở nên rất chua.
Ban ngày, cây sẽ tắt hoạt động của enzyme PEPC và kích hoạt các kênh vận chuyển để đưa axit malic từ không bào ra tế bào chất. Các enzyme khử carboxyl (như enzyme malic) được kích hoạt bởi ánh sáng, phân giải axit malic để giải phóng CO2 ngay bên trong tế bào. Nồng độ CO2 bên trong tế bào tăng vọt, tạo điều kiện lý tưởng cho enzyme RuBisCO hoạt động hiệu quả trong chu trình Calvin (giống hệt cây C3) mà không bị cạnh tranh bởi O2. Lúc này, khí khổng đã đóng chặt để ngăn mất nước.
Các yếu tố điều biến hiệu suất quang hợp
Ánh sáng
Ánh sáng là nguồn năng lượng khởi đầu cho toàn bộ quá trình quang hợp.
- Cường độ ánh sáng: Khi cường độ ánh sáng tăng, hiệu suất quang hợp thường tăng theo cho đến khi đạt đến một điểm gọi là điểm bão hòa ánh sáng. Vượt qua điểm này, cường độ quang hợp không tăng nữa và thậm chí có thể giảm do các sắc tố quang hợp bị tổn thương (quang oxy hóa). Các loài cây khác nhau có điểm bão hòa ánh sáng khác nhau (cây ưa sáng có điểm bão hòa cao hơn cây ưa bóng). Vượt qua điểm này, cường độ ánh sáng cao hơn không làm tăng quang hợp mà có thể gây tổn thương quang học. Điểm bù ánh sáng là cường độ ánh sáng mà tại đó tốc độ quang hợp bằng tốc độ hô hấp.
- Thành phần quang phổ: Quang hợp diễn ra hiệu quả nhất ở vùng ánh sáng đỏ (khoảng 620-750 nm) và vùng ánh sáng xanh tím (khoảng 450-495 nm). Diệp lục hấp thụ kém ở vùng ánh sáng màu xanh lục, đó là lý do tại sao chúng ta thấy lá cây có màu xanh. Tia sáng đỏ thúc đẩy sự hình thành carbohydrate, trong khi tia xanh tím kích thích tổng hợp axit amin và protein.
- Thời gian chiếu sáng: Thời gian chiếu sáng trong ngày càng dài, tổng sản phẩm quang hợp tích lũy được càng nhiều.
Nồng độ CO2
CO2 là nguyên liệu chính cho pha tối (chu trình Calvin) để tổng hợp nên các hợp chất hữu cơ. Nồng độ CO2 trong không khí hiện nay là khoảng 0,04% (400 ppm). Khi nồng độ CO2 tăng, hiệu suất quang hợp sẽ tăng theo. Tuy nhiên, nếu nồng độ tăng quá cao (ví dụ trên 0,2%), cây có thể bị ngộ độc, khí khổng đóng lại và quá trình quang hợp sẽ bị ức chế. Giống như ánh sáng, các loài cây khác nhau cũng có điểm bão hòa CO2 khác nhau.
Nhiệt độ
Nhiệt độ ảnh hưởng trực tiếp đến hoạt động của các enzyme tham gia vào cả pha sáng và pha tối của quang hợp. Mỗi loài thực vật có một khoảng nhiệt độ tối ưu cho quang hợp (thường từ 25-35°C đối với hầu hết các loài cây C3). Nếu nhiệt độ quá thấp (dưới 10°C) hoặc quá cao (trên 40°C), hoạt động của các enzyme sẽ giảm mạnh, làm cho hiệu suất quang hợp giảm hoặc ngừng lại. Nhiệt độ cao còn làm tăng quá trình thoát hơi nước, khiến cây bị mất nước.
Nước
Nước đóng vai trò kép trong quang hợp. Nước là nguồn cung cấp electron (e−) và proton (H+) cho pha sáng thông qua quá trình quang phân ly nước. Nước ảnh hưởng đến độ mở của khí khổng. Khi cây thiếu nước, khí khổng sẽ đóng lại để giảm thoát hơi nước. Điều này đồng thời làm giảm lượng CO2 khuếch tán vào lá, từ đó làm giảm hiệu suất quang hợp. Tình trạng thiếu nước từ 40-60% có thể làm quang hợp giảm mạnh hoặc ngừng hẳn.
Dinh dưỡng khoáng
Các nguyên tố khoáng tuy không phải là thành phần trực tiếp của phản ứng quang hợp nhưng rất cần thiết để duy trì bộ máy quang hợp. Nitơ (N), Phốtpho (P), Lưu huỳnh (S) là thành phần cấu tạo của các enzyme quang hợp. Nitơ (N) và Magie (Mg) là thành phần cốt lõi để tổng hợp nên diệp lục (chlorophyll). Thiếu các nguyên tố này, lá sẽ bị vàng, làm giảm khả năng hấp thụ ánh sáng. Kali (K) tham gia vào việc điều tiết đóng mở khí khổng, ảnh hưởng gián tiếp đến quá trình trao đổi CO2. Mangan (Mn) cần thiết cho phức hệ enzyme quang phân ly nước trong Hệ quang hợp II. Clo (Cl) cần thiết cho quá trình quang phân ly nước và cân bằng thẩm thấu.
Ứng dụng nông học
Các loài cây trồng có đặc điểm quang hợp khác nhau, từ đó quyết định khả năng thích nghi và phân bố không đồng đều trong tự nhiên cũng như trong sản xuất nông nghiệp. Việc nghiên cứu và hiểu rõ đặc điểm quang hợp của từng loài cho phép xác định điều kiện sinh trưởng tối ưu, đồng thời hỗ trợ mở rộng hoặc quy hoạch vùng trồng thích hợp cho các nhóm cây trồng có giá trị kinh tế.
Phân bón cung cấp các nguyên tố khoáng thiết yếu, là thành phần không thể thiếu hoặc là đồng yếu tố (cofactor) cho các phân tử và enzyme tham gia vào quá trình quang hợp. Việc bón phân cân đối và hợp lý sẽ thúc đẩy cây trồng sinh trưởng, phát triển cành lá, ra hoa và đậu quả, từ đó nâng cao năng suất.
Việc bón phân không chỉ đơn thuần là cung cấp thức ăn cho cây. Đó là một sự can thiệp trực tiếp để cung cấp các khối xây dựng nguyên tử cụ thể cần thiết để xây dựng và vận hành bộ máy quang hợp ở hiệu suất cao nhất. Các triệu chứng thiếu dinh dưỡng có thể quan sát được (ví dụ, vàng lá do thiếu N hoặc Mg) là biểu hiện vĩ mô của sự thất bại trong việc tổng hợp diệp lục, phân tử thu nhận ánh sáng.
Thuốc trừ cỏ là các hóa chất được thiết kế để tiêu diệt thực vật không mong muốn, thường bằng cách can thiệp vào các quá trình sinh lý thiết yếu như quang hợp. Các loại thuốc trừ cỏ thành công nhất là những loại nhắm vào các con đường sinh hóa quan trọng đối với thực vật nhưng không tồn tại hoặc khác biệt đáng kể ở động vật. Quang hợp là mục tiêu tối thượng đặc trưng của thực vật. Nhiều loại thuốc trừ cỏ nhắm trực tiếp vào các phản ứng sáng. Các hoạt chất như Atrazine và Diuron liên kết với một protein trong Hệ quang hợp II, chặn dòng chuyền electron. Điều này không chỉ làm ngừng sản xuất ATP và NADPH mà còn dẫn đến sự hình thành các gốc oxy phản ứng cực kỳ phá hoại, phá hủy diệp lục và màng tế bào. Trong vòng vài giờ sau khi phun, quá trình quang hợp chậm lại, và các triệu chứng có thể nhìn thấy như vàng lá và chết mô xuất hiện trong vòng vài ngày.


